Haematoloechus caballeroi Skrjabin & Antipin, 1962
( Figs. 4
View FIGURE 4
& 5
View FIGURE 5
)
Type host: Tlaloc´s leopard frog
Rana tlaloci Hillis & Frost
recorded as Montezuma leopard frog
Rana montezumae
( Caballero, 1942b; Skrjabin & Antipin, 1962).
Type locality: Xochimilco, Mexico City, Mexico ( Caballero, 1942b; Skrjabin & Antipin, 1962).
Site of infection: lungs.
Neotype:
CNHE 10457
.
Paratypes:
CNHE 1428
,
1551
,
1552
,
3376–3379
,
3395
,
3397
,
3399
,
3794
,
4661
,
10458–10460
.
Other hosts and localities: Montezuma leopard frog
R. montezumae, Zempoala
, Morelos (CNHE 10464), Patzcuaro leopard frog
R. dunni Zweifel, Pátzcuaro
, Michoacán (CNHE 10465);
R. dunni, Zacapu
, Michoacán (CNHE 10463).
Other records: Mexico: Tlaloc´s leopard frog
R. tlaloci, Xochimilco
, Mexico City ( Caballero &Sokoloff 1934 as
H. complexus
); Montezuma leopard frog
R. montezumae, Ciénaga de Lerma
, Mexico State (León- Règagnon 1992 as
H. complexus
; Lamothe et al. 1997 as
H. coloradensis Cort, 1915
; León-Règagnon et al. 1999 as
H. complexus
; Pérez-Ponce de León et al. 2000 as
H. coloradensis
and
H. complexus
); big-footed leopard frog
R. megapoda, Cointzio Springs
, Michoacán (Pérez-Ponce de León et al. 2000 as
H. complexus
); Transverse Volcanic leopard frog
R. neovolcanica, Cointzio Springs
, Michoacán (Pérez-Ponce de León et al. 2000 as
H. complexus
); Patzcuaro leopard frog
R. dunni, Pátzcuaro
, Michoacán ( García-Altamirano et al. 1993; Pulido–Flores 1994; Pérez-Ponce de León et al. 2000 as
H. coloradensis
); Lerma Lake salamander
Ambystoma lermaense, Ciénaga de Lerma
, Mexico State (Pérez-Ponce de León et al. 2000 as
H. complexus
; Mata-López et al. 2002 as
H. complexus
).
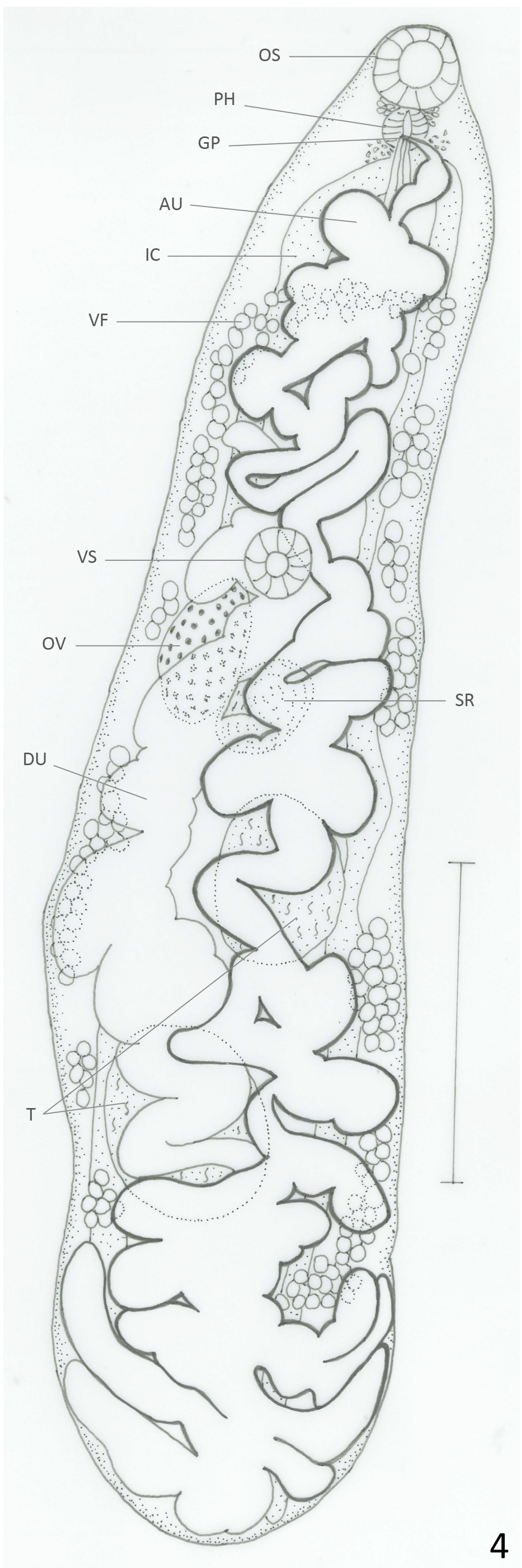
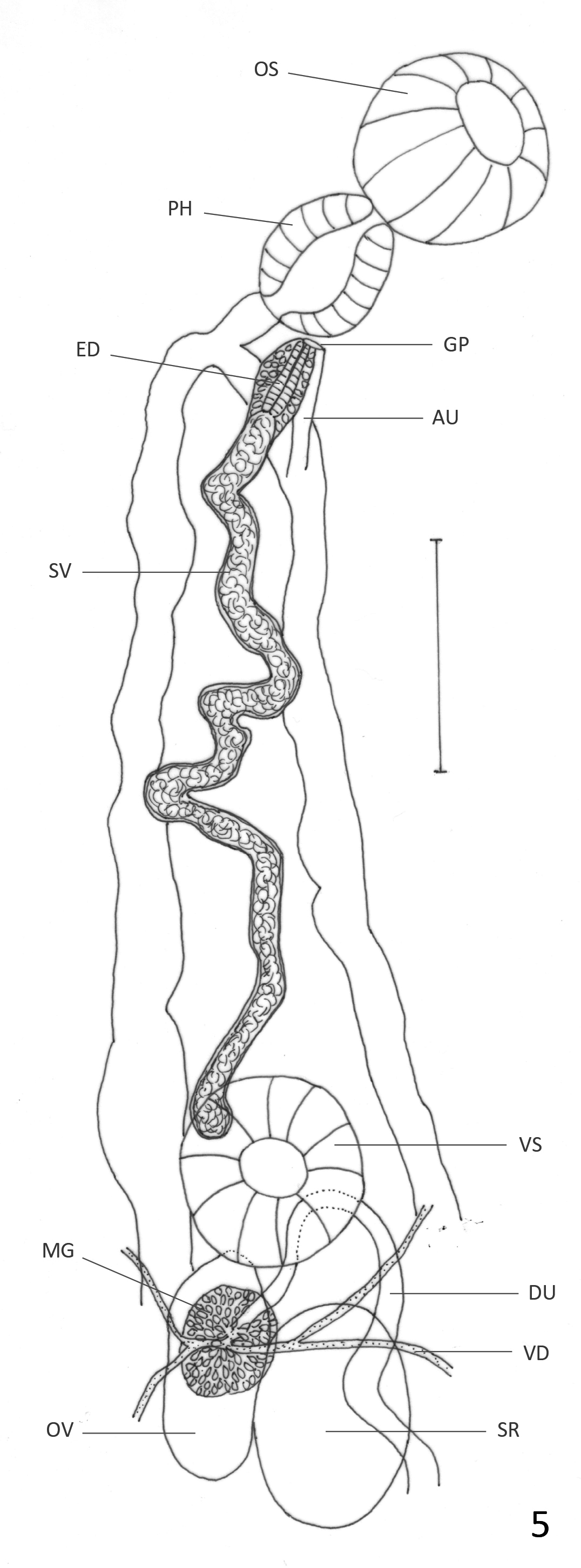
Redescription: Based on 33 mature specimens. Body elongate, with slender anterior region; 2.8–6.6 (5.0) mm long, 0.82–1.88 (1.27) mm of maximum width at testicular region. Tegument covered with thin spines that are larger and more abundant in anterior region; spines easily lost during fixation and staining procedures; 8–16 (13) long. Oral sucker subterminal, round, 243–487 (355) long, 260–511 (387) wide. Pharynx oval, 146–398 (255) long, 130–300 (206) wide; oral sucker/pharynx ratio 1: 0.41–0.86 (0.66). Pharynx and anterior region of esophagus surrounded by gland cells. Esophagus 24–162 (65) long, sometimes obscured by uterus. Ceca bifurcated at 414– 844 (643) from anterior extremity. Ceca terminate blindly near posterior extremity. Ventral sucker round, 219–438 (313) long, 203–430 (311) wide, at 0.98–2.7 (1.9) mm (30%–47% (37.6%) of BL) from anterior extremity. Oral sucker: ventral sucker length ratio 1: 0.53–1.0 (0.81). Testes 2, oval, slightly lobed in some specimens, oblique, inmmediately posterior to ovary. Anterior testis opposite to ovary, 219–795 (477) long, 170–682 (360) wide. Posterior testis 203–852 (557) long, 203–771 (396) wide. Cirrus sac reach anterior border of ventral sucker, mostly obscured by ascending uterus; internal seminal vesicle, elongate, slightly coiled. Ejaculatory duct strongly muscular, 190–200 (195) long, surrounded by prostatic gland cells. Ovary oval, 243–665 (414) long, 203–763 (313) wide; at 1.1–2.9 (2.1) mm (33%–51% (42%) of BL) from anterior extremity. Seminal receptacle adjacent partially overlapped with ovary; 203–730 (412) long, 138–568 (300) wide. Mehlis gland dorsal to seminal receptacle. Laurer’s canal not observed. Vitellaria in clusters overlapped with each other, distributed laterally, dorsally invade space between ceca in their anterior limit and in post-testicular region. Anterior limit of distribution 390–1607 (1055) (6.5%–28% (21%) of BL) from anterior end. Follicles extend to halfway between posterior testis and posterior end; in some specimens they extend to level of posterior testis on ovarian side of body, and more posteriorly on side opposite to ovary. Descending part of uterus form transverse and diagonal loops on ovarian side of body, partially overlapped with testis, filling intra- and extracecal space towards posterior end of body. Ascending uterus form one or two short diagonal loops oriented anteriorly on each side of body, and continues with transverse or diagonal loops that occasionally invade both sides of body, not totally overlapped with testes or ovary, and fill with transverse loops entire preovarian region. Genital pore median, ventral to pharynx. Eggs dark brown, 31–40 (36) long, 17–23 (20) wide. Excretory vesicle not observed. Excretory pore terminal.
Remarks: In their revision of the family
Plagiorchiidae Lühe, 1901
, Skrjabin & Antipin (1962) described
Haematoloechus caballeroi
based on a specimen that Caballero (1942b) collected from Tlaloc´s leopard frog
R. tlaloci in Xochimilco
, Mexico, and was originally identified as
H. complexus
. This description was based on a single specimen, and was not recognized by any later author; all subsequent records of that morphotype from Mexico were assigned to
H. complexus
or
H. coloradensis
. Recent molecular and morphological evidence has shown that
H. complexus
sensu stricto does not occur in Mexico, but that there is a complex of species closely related to
H. complexus
, most undescribed ( León-Règagnon & Brooks 2003; Bolek & Janovy 2007a; León- Règagnon 2010; León-Règagnon & Romero–Mayén 2017). The present species is one of those (GenBank
AF532138
View Materials
, León-Règagnon & Brooks 2003).
Haematoloechus caballeroi
differs from most other species in the genus, but resembles the following 19 species by lacking longitudinal uterine loops that reach the posterior testis:
H. arequipensis Ibáñez & Córdoba, 1979
,
H. aubriae Bourgat, Roure & Kulo, 1996
,
H. coloradensis
,
H. complexus
,
H. confusus Ingles, 1932
,
H. danbrooksi
,
H. dollfusinum ( Odening, 1958)
,
H. elongatus Caballero & Sokoloff, 1934
,
H. fuelleborni ( Travassos & Darriba, 1930)
,
H. humboldtensis
,
H. illimis
,
H. kernensis Ingles, 1932
,
H. longicollum
,
H. medioplexus
,
H. meridionalis
,
H. oxyorchis Ingles, 1932
,
H. parcivitellarius Caballero, 1942
,
H. pukinensis Ibáñez & Córdoba, 1979
, and
H. pulcher
. It differs from
H. aubriae
,
H. danbrooksi
,
H. medioplexus
, and
H. meridionalis
in the large size of the ventral sucker compared to the oral sucker, which is less than one third in those four species vs more than half in
H. caballeroi
( Table 2) ( Stafford 1902; Bourgat et al. 1996; León-Règagnon et al. 2001; León- Règagnon & Paredes-Calderón 2002).
Haematoloechus coloradensis
,
H. confusus
, and
H. oxyorchis
differ from
H. caballeroi
in the arrangement of the uterine loops, which are strictly intercecal in those species ( Cort 1915; Ingles 1932; Bolek & Janovy 2007a), while they invade the extracecal region in
H. caballeroi
.
Haematoloechus caballeroi
differs from
H. arequipensis
in having oval rather than lobed testes ( Ibañez & Córdoba 1979). It differs from
H. illimis
and
H. dollfusinum
in the shape of the ovary, which is lobed in those species ( Caballero 1942a) and oval in
H. caballeroi
. The presence of diagonal uterine loops directed anteriorly at the posterior end of the body differentiates
H. caballeroi
from
H. humboldtensis
,
H. longicollum
,
H. parcivitellarius
, and
H. pulcher
, in which the diagonal uterine loops are either absent or directed posteriorly ( Caballero 1942b; Bravo–Hollis 1943; Zamparo et al. 2011; León-Règagnon & Romero–Mayén 2017).
Haematoloechus caballeroi
most closely resembles
H. complexus
,
H. elongatus
,
H. fuelleborni
,
H. kernensis
and
H. pukinensis
. It differs from
H. fuelleborni
,
H. complexus
and
H. elongatus
in having a larger pharynx and ventral sucker compared to the oral sucker (1:0.45, 1:0.56, 1:51 & 1:0.5, 1:0.71, 1:0.70 respectively vs 1:0.66 & 1: 0.81 in
H. caballeroi
) ( Travassos & Darriba 1930; Caballero & Sokoloff 1934; Bolek & Janovy 2007a). It also differs from
H. fuelleborni
in the distribution of the vitellaria; while they are limited to two groups, one anterior to the ventral sucker and other posterior to the testes in the South American species, they are distributed continuously from the anterior region of the ventral sucker to the posterior region of the testes in
H. caballeroi
. It also differs from
H. complexus
in the distribution of the vitellaria which are distributed asymmetrically in that species, being more restricted to the ovarian side of the body ( Bolek & Janovy 2007a), while in
H. caballeroi
they reach the posterior region of the testes on both sides of the body. In addition to the differing size of the ventral sucker and pharynx,
H. elongatus
differs from
H. caballeroi
in body size, which is much larger in that species ( 9.5 mm vs 5.1 mm) than in
H. caballeroi
.
Haematoloechus caballeroi
differs from
H. kernensis
in the size of the ventral sucker compared with the oral sucker, which is larger in that species (1:1, vs 1: 0.82 in
H. caballeroi
). It also differs from
H. kernensis
in the distribution of the vitellaria, which do not invade the intercecal region in the post-testicular region in that species, and in the arrangement of the uterus, which forms a few diagonal loops in the pre-acetabular and post-testicular region in
H. kernensis
, while it is filled with transverse and diagonal loops in both areas in
H. caballeroi ( Ingles 1932)
.
Haematoloechus caballeroi
differs from
H. pukinensis
in the arrangement of the uterine loops; while in
H. pukinensis
the ascending part of the uterus forms a few transverse loops in the pre-acetabular region ( Ibáñez & Córdoba 1979), in
H. caballeroi
the ascending uterus entirely fills the pre-acetabular region. The distribution of the vitellaria is also different in
H. pukinensis
, being more restricted in the ovarian side of the body in that species.
......continued on the next page ......continued on the next page
H = Hasegawa et al., 2013.
L = León-Règagnon 2010.
LB = León-Règagnon & Brooks 2003.
LGA = León-Règagnon et al. 2005.
RLP = Razo-Mendívil et al. 2004, 2006.
T = Tkach et al. 2000.
Z = Zikmundova et al. 2014.
1 originally recorded as
H. coloradensis
.
2 originally recorded as
H. cf. complexus
.
3 originally recorded as
H. varioplexus
.