
Grubeulepis kurnai, Woolley, Skipton & Wilson, Robin S., 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.205570 |
|
DOI |
https://doi.org/10.5281/zenodo.5681725 |
|
persistent identifier |
https://treatment.plazi.org/id/03A187B0-720C-4D6B-FF2E-F9C2FD3ECFA0 |
|
treatment provided by |
Plazi |
|
scientific name |
Grubeulepis kurnai |
| status |
sp. nov. |
Grubeulepis kurnai View in CoL sp. nov.
Figures 1–7
Material Examined. Holotype: Australia: Bass Strait: near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 5 S8, 38 º22´S 147º12´E, 28 Jan 1990, 15.3 m, MV F64546 View Materials . Sampling method, 0.25 m2 quadrat driven 10 cm into sand; enclosed 10 cm sediment removed by water venturi suction and filtered directly through a 750 micron mesh bag on bottom (sampling method identical for all MSL-LV stations below).
Paratypes (6 specimens): Australia: Bass Strait: 1 km off Delray Beach, Stn MSL-LV 5 D2, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 1 paratype, MV F64563 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 5 S6, 38 º22´S 147º12´E, 28 Jan 1990, 16 m, 1 paratype, MV F64569 View Materials ; 1 km off Delray Beach, Stn MSL-LV 5 T5, 38 º14´S 147º22´E, 29 Jan 1990, 15 m, 1 paratype, MV F64550 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 4 S5, 38 º22´S 147º12´E, 23 Jan 1989, 15.5 m, 1 paratype, MV F64551 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 4 S6, 38 º22´S 147º12´E, 23 Jan 1989, 16 m, 1 paratype, MV F64561 View Materials ; 1 km off Delray Beach, Stn MSL-LV 1 D8, 38 º14´S 147º22´E, 23 Jan 1989, 16.5 m, 1 paratype, MV F64579 View Materials .
Non-type material: Australia: Bass Strait: 1 km off Delray Beach, Stn MSL-LV 3 T2, 38 º14´S 147º22´E, 27 Jan 1989, 16.5 m, 1 specimen, MV F64554 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 D2, 38 º14´S 147º22´E, 17 Jan 1989, 16 m, 1 specimen, MV F64568 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 4 S8, 38 º22´S 147º12´E, 23 Jan 1989, 15.3 m, 1 specimen, MV F64567 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 5 S7, 38 º22´S 147º12´E, 28 Jan 1990, 16 m, 1 specimen, MV F64566 View Materials ; 1 km off Delray Beach, Stn MSL-LV 6 D8, 38 º14´S 147º22´E, 29 Jan 1990, 16.5 m, 1 specimen, MV F64562 View Materials ; 1 km off Delray Beach, Stn MSL-LV 5 D4, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 1 specimen, MV F64559 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 D5, 38 º14´S 147º22´E, 17 Jan 1989, 16 m, 2, MV F64558 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 D3, 38 º14´S 147º22´E, 17 Jan 1989, 16 m, 1 specimen, MV F64564 View Materials ; 1 km off Delray Beach, Stn MSL-LV 3 D7, 38 º14´S 147º22´E, 11 Jan 1989, 15.5 m, 1 specimen, MV F64557 View Materials ; 1 km off Delray Beach, Stn MSL-LV 3 T3, 38 º14´S 147º22´E, 24 Jan 1989, 14.8 m, 5 specimens, MV F64555 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 T2, 38 º14´S 147º22´E, 24 Jan 1989, 16.5 m, 1 specimen, MV F64541 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 5 S4, 38 º22´S 147º12´E, 28 Jan 1990, 14.5 m, 2 specimens, MV F64547 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 3 S1, 38 º22´S 147º12´E, 23 Jan 1989, 15.5 m, 1 specimen, MV F64581 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 4 S1, 38 º22´S 147º12´E, 23 Jan 1989, 15.5 m, 1 specimen, MV F64544 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 5 S5, 38 º22´S 147º12´E, 28 Jan 1990, 15.5 m, 1 specimen, MV F64542 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 6 S4, 38 º22´S 147º12´E, 28 Jan 1989, 14.5 m, 1 specimen, MV F64540 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 T1, 38 º14´S 147º22´E, 24 Jan 1989, 16.6 m, 1 specimen, MV F64548 View Materials ; 1 km off Delray Beach, Stn MSL-LV 5 T8, 38 º14´S 147º22´E, 29 Jan 1990, 15.4 m, 3 specimens, MV F64538 View Materials ; 1 km off Delray Beach, Stn MSL-LV 6 D3, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 3 specimens, MV F64549 View Materials ; 1 km off Delray Beach, Stn MSL-LV 5 D2, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 1 specimen, MV F64582 View Materials ; 1 km off Delray Beach, Stn MSL-LV 6 D5, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 1 specimen, MV F64571 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 D7, 38 º14´S 147º22´E, 11 Jan 1989, 15.5 m, 1 specimen, MV F64536 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 6 S6, 38 º22´S 147º12´E, 28 Jan 1990, 16 m, 1 specimen, MV F64578 View Materials ; Grubeulepis sp MoV 737; 500 m off Woodside Beach, eastern Bass Strait, Stn SWOP 93 18 3, 38º33.05´S 146º59.317´E, 1993, 7 m, 1 specimen, MV F166648; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 4 S8, 38 º22´S 147º12´E, 23 Jan 1989, 15.3 m, 1 specimen, MV F64576 View Materials ; 1 km off Delray Beach, Stn MSL-LV 2 T2, 38 º14´S 147º22´E, 1 Jan 1989, 16.5 m, 2 specimens, MV F64553 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 3 S7, 38 º22´S 147º12´E, 23 Jan 1989, 16 m, 1 specimen, MV F64570 View Materials ; 1 km off Delray Beach, Stn MSL-LV 6 D2, 38 º14´S 147º22´E, 29 Jan 1990, 16 m, 2 specimens, MV F64556 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 6 S5, 38 º22´S 147º12´E, 28 Jan 1990, 15.5 m, 2 specimens, MV F64545 View Materials ; 1 km off Delray Beach, Stn MSL-LV 4 D6, 38 º14´S 147º22´E, 17 Jan 1989, 15.8 m, 3 specimens, MV F64552 View Materials ; 1 km off Delray Beach, Stn MSL-LV 3 D5, 38 º14´S 147º22´E, 17 Jan 1989, 16 m, 1 specimen, MV F64574 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 1 S5, 38 º22´S 147º12´E, 9 Jan 1989, 15.5 m, 1 specimen, MV F64577 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 3 S6, 38 º22´S 147º12´E, 23 Jan 1989, 16 m, 1 specimen, MV F64580 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 2 S6, 38 º22´S 147º12´E, 9 Jan 1989, 16 m, 1 specimen, MV F64583 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 1 S2, 38 º22´S 147º12´E, 9 Jan 1989, 16 m, 2 specimens, MV F64575 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 1 S4, 38 º22´S 147º12´E, 9 Jan 1989, 14.5 m, 1 specimen, MV F64573 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 2 S2, 38 º22´S 147º12´E, 9 Jan 1989, 16 m, 2 specimens, MV F64572 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 1 S6, 38 º22´S 147º12´E, 9 Jan 1989, 16 m, 1 specimen, MV F64565 View Materials ; 1 km off Delray Beach, Stn MSL-LV 2 D6, 38 º14´S 147º22´E, 23 Jan 1989, 15.8 m, 1 specimen, MV F64560 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 2 S5, 38 º22´S 147º12´E, 9 Jan 1989, 15.5 m, 1 specimen, MV F64543 View Materials ; near Seaspray, 1 km off The Honeysuckles, Stn MSL-LV 1 S7, 38 º22´S 147º12´E, 9 Jan 1989, 16 m, 2 specimens, MV F64537 View Materials ; 1 km off Delray Beach, Stn MSL-LV 1 T4, 38 º14´S 147º22´E, 1 Jan 1989, 14.8 m, 1 specimen, MV F64539 View Materials .
Description. Adults: Size range of material examined 3–24 mm long (n= 42; holotype 23 mm), 1–7 mm wide including chaetae (holotype 6 mm), 20–37 segments (holotype 37 segments). Elytrae 12 pairs on segments 2, 4, 5, 7, 9, 11, 13, 15, 17, 19, 21 and 24, becoming more elongate towards the posterior end (Figure 1A). First pair of elytrae with 0–10 papillae on the anterior margin (6 papillae in the holotype), elytrae 2 to 12 with distinct lateral leaf projection between the anterior and posterior surfaces. Elytron 2 with 4–23 lateral lobes (23 in holotype). Elytron 8 with 7–11 lateral lobes (9 in holotype) and elytron 12 with 17–27 (17 in holotype). Lateral processes of elytrae digitiform, smaller individuals tend to have articulate lateral processes (Figures 1B–E).
FIGURE 1. Grubeulepis kurnai sp. nov., holotype MV F64546 View Materials : A, Entire animal, dorsal view, first right elytron removed; B, 1st right elytron; C, 2nd left elytron; D, 8th left elytron; E, 12th left elytron.
Branchiae 11 pairs on segments 8, 10, 12, 14, 16, 18, 20, 22, 23, 25, 26 as defined by the presence of distally attached branchial cirrus, replaced by fleshy lamellae on the 27th and following segments. The lamellae are foliform, reducing in size towards the posterior end ( Figure 3 View FIGURE 3 D). The branchiae project from the dorsal region of the parapodia and have dorsal and ventral ciliated bands, along with distally attached branchial cirrus ( Figure 3 View FIGURE 3 B).
Prostomium ( Figure 2 View FIGURE 2 A) is covered by the 2nd segment, antenna and palps reduced and eyes absent. Median antenna small and aspidate, attached to the prostomium on the dorsal margin. Lateral antennae conical and attach laterally relative to the median antenna.
Parapodia biramous, each ramus supported by single amber aciculum, notoaciculum of median and posterior segments with hooked tip, neuroaciculum throughout expanded distally to form malleiform distal plates ( Figure 3 View FIGURE 3 A–D). Dorsal-most notochaetae amber, curved with the distal edge serrated and disciform tip ( Figure 4 View FIGURE 4 A). Ventral notochaetae are also curved but not serrated, and form a sharp point at the tip. Notopodial capillaries densely bunched on the lower half of the notopodium, projecting from a fleshy lump on the lower notopodial lobe. Capillaries of two forms, the first entirely smooth and tapering to a fine point ( Figure 4 View FIGURE 4 B), the second with serrated ventral margin continuing for a third of the length of the capillary ( Figure 4 View FIGURE 4 C). Dorsal-most neurochaetae are pectinate ( Figure 4 View FIGURE 4 F), more ventral neurochaetae are smooth capillaries that taper to a fine point, capillary chaetae becoming thinner in more ventral positions ( Figure 4 View FIGURE 4 D, E). On posterior segments upper notochaetae and neurochaetae are both much stouter, notochaetae are sharply bent, serrated on the outer margin and taper evenly to a point ( Figure 4 View FIGURE 4 H), neurochaetae are longer, sharply bent with a distinct corrugated pattern on the outer margin distal to the bend and narrow abruptly to a point ( Figure 4 View FIGURE 4 G).
Pygidium of all intact specimens with right cirrus long, minutely papillated on one side only for most of the length except the basal-most part; left cirrus ovoid ( Figure 2 View FIGURE 2 B).
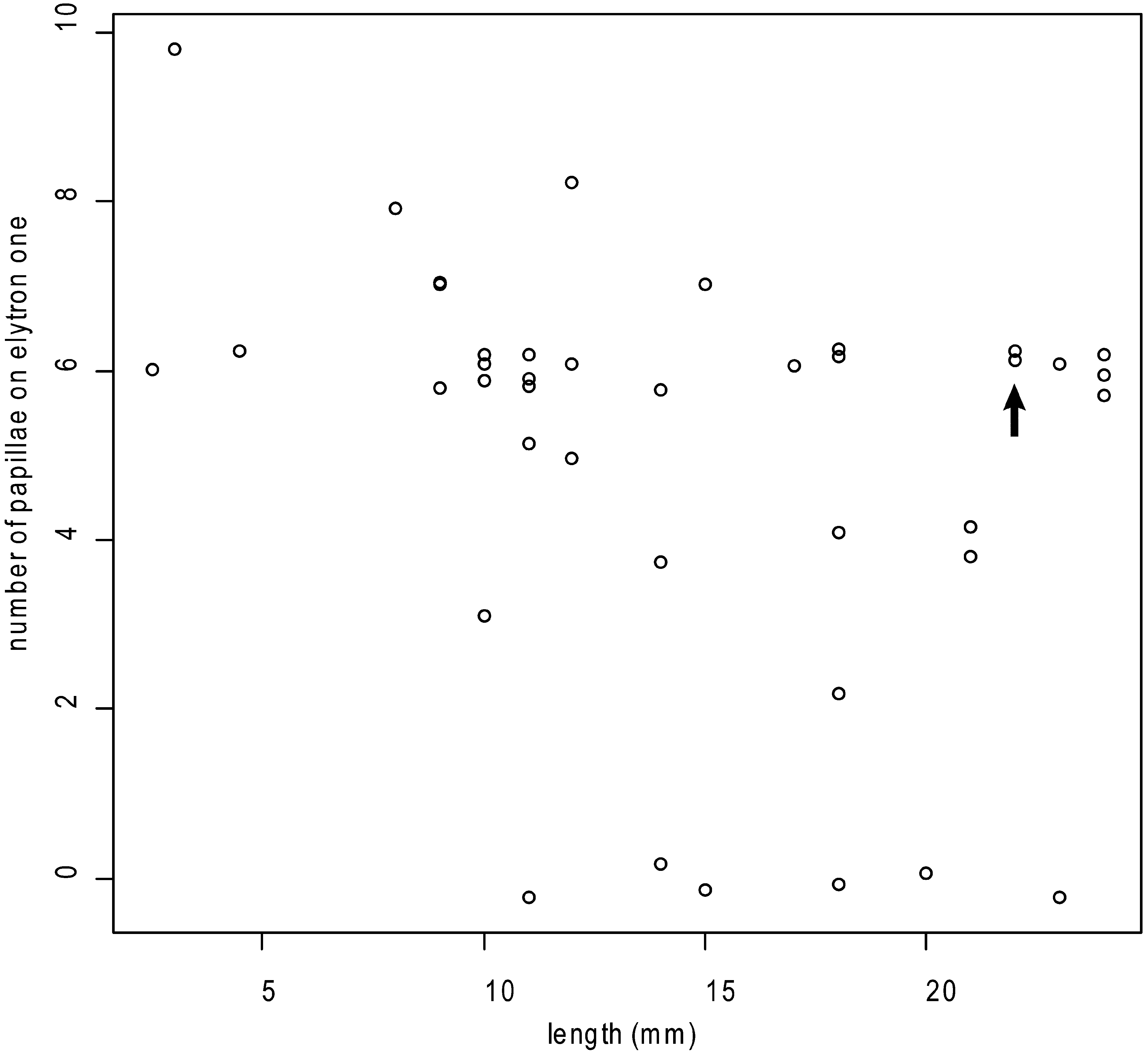
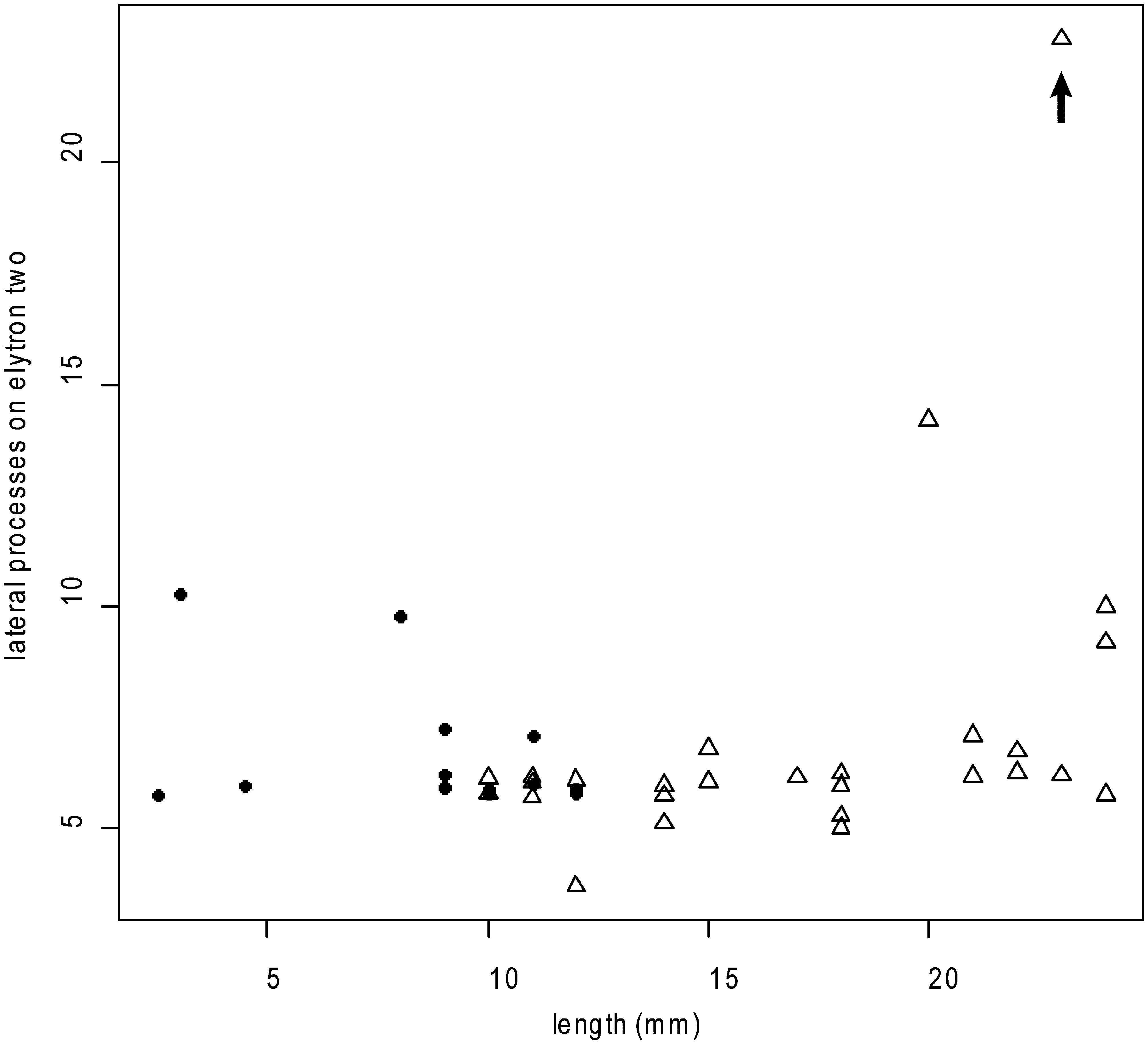
Juveniles: Size range of material examined 1.5– 7 mm long (n= 11; holotype 5 mm), 1–2.5 mm wide (holotype 2 mm) including chaetae, 12–26 segments (holotype 25). Elytrae 12 pairs on segments 2, 4, 5, 7, 9, 11, 13, 15, 17, 19, 21 and 24, however the 11th and 12th pairs are greatly reduced and the 12th pair of elytrae is absent in some of the smaller juveniles. First pair of elytrae have 1–11 papillae on the anterior margin (8 in holotype); a scatter plot of number of papillae on the anterior margin of the first elytron versus body length ( Figure 6 View FIGURE 6 ) shows that size is a poor predictor of number of papillae (R2= 0.1). Elytrae 2 to 12 show the same lateral anterior to posterior projections, however the biggest difference seen between juveniles and adults is the presence of articulate lateral processes on the elytrae of juveniles (Figures 5E–G). A scatterplot of number of lateral processes on the second elytron versus body length ( Figure 7 View FIGURE 7 ) demonstrates the progression from articulate to inarticulate lateral processes with increasing size of specimen.
Branchiae display the same morphology and attach from the eighth segment onwards, however due to the reduced size of juveniles the posterior branchiae are either not developed or absent, this is evident in the holotype with eight pairs of branchiae.
Biramous parapodia similar in form to adults (Figures 5A–D). Notopodial chaetae similar to that of adults; hooked notochaetae include forms with smooth and forms finely serrated on distal margin. Notopodial capillaries of juveniles are equivalent to those seen in adults as are upper pectinate, and smooth fine ventral neurochaetae. However the dorsal neurochaetae of juveniles differ from adults: in the anterior and mid-regional segments neurochaetae of juveniles are stout and slightly denticled, compared to smooth slender chaetae ending in a fine point which are seen in adult animals. Posterior stout hooked neurochaetae from the dorsal region of the neuropodia also differ between juveniles and adults: juveniles have two spinous rows on the distal edge (Figure 5H), whereas adults have a distinctly corrugated pattern ( Figure 4 View FIGURE 4 F).
Other morphological characters showed no differences between juvenile and adult specimens.
Etymology. The species name kurnai pays respect to the Aboriginal people of the Kurnai Nation whose traditional lands in eastern Victoria include that part of the Bass Strait coast where Grubeulepis kurnai sp. nov. occurs.
Distribution and habitat. Grubeulepis kurnai sp. nov. occurs in eastern Bass Strait, southeastern Australia in depths of 7–16.6 metres from medium to coarse sand sediments with mean grain size in the range 0.3–2.1 mm; mud content, measured at 6 stations, was in the range 0.7–4.7%. Grubeulepis kurnai sp. nov. was only collected using a SCUBA-operated airlift; extensive sampling with epibenthic sleds and Smith McIntyre Grabs throughout Bass Strait, including many adjacent stations and mostly in the depth range 22–90 m ( Wilson & Poore, 1987), failed to collect even a single eulepethid, nor were any found during two extensive surveys of nearby Port Phillip Bay using Smith McIntyre Grabs in depths 5–22 m ( Poore, et al., 1975; Wilson, et al., 1998). One possible explanation is that Grubeulepis kurnai sp. nov. lives in tubes of another benthic organism and is not normally collected unless the suction of SCUBA-collected airlift is applied. The habitat at one location was described by the collector thus: “Fine-medium grained sandy bed. Very sparse epibiota, typically a few red algae, some Pseudogorgia godeffroyi , drift from scattered inshore reef comprising of Amathia , sometimes with pycnogonid passengers and the small gastropods. Strong tidal longshore currents with rippled bed.” (J.E.Watson, pers. comm. 2010).
Discussion. The genus Grubeulepis contains ten species ( Pettibone 1986, Nishi 2001), which are compared in Table 2 View TABLE 2 .
Grubeulepis kurnai sp. nov. is most similar to G. g e a y i ( Fauvel, 1918), but can be separated by the differences in the number of branchiae (11 pairs in G. k u r n a i versus 13 pairs in G. g e a y i), the number of papillae on elytrae (0– 10 in the first elytron in G. kurnai versus 3–4 in G. geayi ), the number of lateral processes of elytrae (in 2nd, 8th and 12th of both species, see Table 2 View TABLE 2 ), the shape of posterior lamellae (foliform in G. k u r n a i versus subconical in G. geayi ), the notochaetae and neurochaetae shape (see Table 2 View TABLE 2 ), and the longer anal cirrus (minutely papillated on one side in G. k u r n a i versus smooth in G. g e a y i). The two species also differ the segments where the posterior lamellae start to form although of course this character is dependent on number of branchiae, which they replace posteriorly, thus posterior lamellae commence on segment 27 in G. kurnai sp. nov. and on segment 28 in G. g e a y i. The shape of the posterior lamellae differs in G. kurnai sp. nov. with leaf shaped (foliform), compared to G. g e a y i in which the posterior lamellae are subreniform and subconical.
Additional differences between G. k u r n a i sp. nov. and G. g e a y i include variation in the notochaetae and the neurochaetae; a more spinous and disciform tipped hooked notochaetae is present in G. k u r n a i sp. nov. ( Figure 4 View FIGURE 4 A). Grubeulepis kurnai sp. nov. lacks the limbate chaetae present in G. g e a y i ( Pettibone, 1969: Figure 18e, c). The upper neurochaetae of posterior segments in G. k u r n a i sp. nov. are very stout, curved downwards and has a distinct corrugation on distal edge ( Figure 4 View FIGURE 4 G) whereas in G. g e a y i the corresponding neurochaetae of posterior segments are stout, taper abruptly to long fine tips and are smooth ( Pettibone, 1969: Figure 20b). (Stout curved distally corrugated upper neurochaetae similar to those in G. k u r n a i sp. nov. are also present on posterior segments in G.mexicana ; see Pettibone 1969: Figure 27c).
FIGURE 5. Grubeulepis kurnai sp. nov., paratype F64579 View Materials , juvenile: A, paradodium segment 3; B, branchial parapodium midbody; C, elytigerous parapodium, mid-body, D, elytrigerous parapodium, posterior segment; E, 1st elytron; F, 2nd elytron; G, 8th elytron; H, dorsal neurochaeta, segment 21; I, dorsal hooked notochaeta, segment 12; J, ventral hooked notochaeta, segment 12. E–G share scale bar; H–J share scale bar.
Other apparent differences between G. kurnai sp. nov. and G. g e a y i must be interpreted cautiously since they are size-related: papillae are more numerous on the first elytrae in G. k u r n a i sp. nov. compared to G. g e a y i, however as demonstrated above the number of papillae on the first elytrae decreases with size ( Figure 6 View FIGURE 6 ). Grubeulepis kurnai sp. nov. has more numerous lateral processes on 2nd, 8th and 12th elytrae in comparison to G. g e a y i (see Table 2 View TABLE 2 ). However, in our material articulate lateral processes occur in smaller specimens, and are apparently replaced by non-articulate lateral processes in larger specimens ( Figure 7 View FIGURE 7 ). Pettibone (1986) notes a similar trend in the presence of articulate lateral processes in juveniles to non-articulate in adult specimens in G. mexicana . We have provided this information in Table 2 View TABLE 2 based on published information but it is not clear whether descriptions have been based on juveniles or adults, therefore caution is required when using form of lateral processes on eyltrae for taxonomic purposes.
Grubeulepis kurnai sp. nov. is also similar to G. mexicana , G. katzmanni and G.t e b b l e i in that all have 11 pairs of branchiae. However Grubeulepis mexicana differs from G. kurnai sp. nov. in that lateral processes of elytrae are biarticulate and tripartite in G. mexicana , and not articulate in G. kurnai sp. nov. in all specimens exceeding 12 mm in length. Grubeulepis kurnai sp. nov. lacks eyes, and additional differences separating the new species from Grubeulepis katzmanni and G. tebblei include form of posterior lamellae, form of notochaetae, presence of limbate neurochaetae and form of the anal cirrus ( Table 2 View TABLE 2 ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |