
Gerstfeldtiancylus ushunensis, Shirokaya, Alena, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.176562 |
|
DOI |
https://doi.org/10.5281/zenodo.5618836 |
|
persistent identifier |
https://treatment.plazi.org/id/B35D667A-002D-B303-9DD2-F93EFAD6DFCF |
|
treatment provided by |
Plazi |
|
scientific name |
Gerstfeldtiancylus ushunensis |
| status |
sp. nov. |
Gerstfeldtiancylus ushunensis View in CoL sp. nov.
( Figs 3–7 View FIGURE 3 View FIGURE 4 View FIGURE 5. A – B View FIGURE 6 View FIGURE 7 )
Type material: Sixteen specimens from the type locality. Holotype: Russia, Lake Baikal, Malye Olkhonskie Vorota, Ushun Bay, 53°00’54’’ N 106°56’96’’ E, 20–22 m depth, on stones, coll. I.V. Khanaev and V.F. Skudenko 0 7.07.2003 ( ZIN RAS 1/521–2006). Paratypes: 4–22 m depth ( ZIN RAS 2/521–2006, 3/521–2006; LIN SB RAS 904, 905).
Etymology. The species is named after its type locality — Ushun Bay.
Material examined. Teleoconch: holotype ( ZIN RAS 1/521–2006) and 15 paratypes ( ZIN RAS 2/521– 2006, 3/521–2006, LIN SB RAS 904, 905); alimentary system, odontophore and jaw: holotype and ten paratypes ( ZIN RAS 2/521–2006, 3/521–2006); protoconch: four paratypes (LIN SB RAS 904, 905); radula and male copulatory organ: five paratypes ( ZIN RAS 2/521–2006); mantle and shell adductor muscles: seven paratypes ( ZIN RAS 2/521–2006). Fourteen syncapsulae were measured.
Description. Shell ( Fig. 3 View FIGURE 3 A–D): Large (aperture up to 11 mm long), relatively high (in mature specimens height and aperture length ratio is 0.34–0.43). Apex slightly displaced to the left (distance from apex to the left aperture edge is 0.76–0.99 of the distance to the right one) and distant from the anterior edge of aperture by 0.58–0.68 of its length (in projection to longitudinal axis of aperture). Distance from apex to the frontal aperture edge along a chord of the anterior (maximal) slope is 0.69–0.77. All four shell slopes slightly convex. Aperture broad oval, its width and length ratio is 0.79–0.84. Protoconch cap-shaped, relatively low (H/h = 8.5); its apical part bears two tubercles, the slopes underneath are slightly convex ( Fig. 3 View FIGURE 3 E–G). Protoconch surface has a distinct reticular microsculpture formed by transecting radial costae and concentric cords and distinct rectangular grooves 0.020–0.027 mm long and 0.014–0.020 mm wide ( Fig. 3 View FIGURE 3 K). Reticular sculpture gradually turns in a concentric one in the direction from the apical part to the protoconch edge ( Fig. 3 View FIGURE 3 I–J). Initial plate lacks any sculpture ( Fig. 3 View FIGURE 3 H), its diameter being about 0.12 mm and forms an angle of 13–14° to the protoconch aperture plane. Protoconch forms an angle of 13–14° to the plane of teleoconch aperture. Dimensions of holotype protoconch: length (l) = 2.44 mm, width (w) = 1.70 mm, height (h) = 0.35 mm. Dimensions of holotype and paratype teleoconchs are given in Table 1 View TABLE 1 .
Radula ( Fig. 4 View FIGURE 4 ) of mature specimens: 74–90 transverse teeth rows. Each row has 57–65 teeth: 16–23 lateral and 7–15 marginal teeth on each side of the central tooth. Central tooth has a narrow base and a short cutting edge with one median cusp and 3–4 wrinkles on both sides. Lateral teeth have a narrow long cutting edge with two terminal cusps. Inner marginal teeth have a broad base and short cutting edge (cusps not developed). The remaining marginal teeth have no cutting edge.
Alimentary system ( Fig. 5A View FIGURE 5. A – B , C–H): Jaw horseshoe-shaped, consists of 76–101 lanceolate scales. Each lateral part of the jaw has up to 32–43 scales, dorsal — 12–15. Buccal bulb is rounded, its length making about 1/3 of the body length. Odontophore of living specimens is red. Salivary glands are short, slender (about 0.1 mm in diameter), proximal section expanded to 0.25–0.3 mm in diameter. Radular sac is short and its length does not exceed 1/6 of the body length. Stomach caecum is pyriform, branching dorso-laterally from the stomach and directed to the left. Dorsal digestive gland pore larger than ventral one. First intestinal loop is positioned on the right side of the body, the second one in the left posterior quarter of it. Digestive gland is large and occupies almost 2/3 of the visceral mass.
Shell adductor muscles ( Fig. 5B View FIGURE 5. A – B ): Left anterior adductor is slender with an elongated surface directed perpendicular to the longitudinal axis of the body or curved like an arch. Right anterior adductor consists of two parts. One is located closer to the anterior body end; it has an arch-shaped surface and is positioned on one line with the left anterior adductor. Other one with a broad oval surface is parallel to the longitudinal axis of the body. Left and right adductors are almost equal in height. Right half of posterior (original) adductor is substantially elongated and anteriorly directed. Its surface is arch-shaped. Surface area of posterior adductor is 2– 2.5 times as large as that of each anterior adductor.
Male copulatory organ ( Fig. 6 View FIGURE 6 F–G): Flagellum nearly as long as the total length of penis sheath and preputium. In cross-section flagellum is even all the way through and the distal end is slightly inflated. Penis sheath is long, cylindrical, evenly wide. Penis clavate, having a terminally located deferent duct opening. Penis length is 3/5 of the sheath length. Proximal part of penis is twice as wide as the distal one. Preputium is cylindrical and it forms a globose chamber with muscular walls in the proximal part. The ratio of the chamber width to the preputium width is 1.2–1.3. Sarcobellum and velum are well developed. Preputium is as long as the penis sheath. Pilasters are pronounced along the whole preputium.
Egg masses ( Fig. 7 View FIGURE 7 A–C, Table 2 View TABLE 2 ): New syncapsulae are bluish-translucent with convex upper surface, large (l = 4.8–7.0 mm, w = 4.0– 6.5 mm, h = 1.3–2.5 mm). The clutch is oval or ellipsoidal. Extrasyncapsular tunic is very broad, with uneven fimbriated margins. Terminal part of the clutch protrudes like a spine. One syncapsula contains two to five egg capsulae. Egg capsulae are very large (up to 2.8 mm long and 2.3 mm wide), oval with centrally or peripherally positioned eggs. Unrounded stones free of plant coatings serve as substratum.
Differential diagnosis. By shell morphology Gerstfeldtiancylus ushunensis is most similar to G. ro e p - storfi. The teleoconchs of both species are relatively low (H/L = 0.30–0.47) and the apex is distant from the anterior edge of aperture by little over 1/2 of its length. G. ro e p s t o r f i differs by an apex location precisely on the longitudinal axis of the shell. The shells of G. kotyensis and G. re n a rd i i are higher (H/L = 0.45–0.60) and the apex is more distant from the anterior edge of the aperture (3/4 of its length). The apex of G. kotyensis is located on the longitudinal shell axis, that of G. re n a rd i i is considerably shifted to the left. In contrast to G. ushunensis , all remaining species of the subgenus have a concave or straight posterior teleoconch slope and a relatively high protoconch (H/h = 4–5). Radular morphology of G. ushunensis and G. re n a rd i i is very similar.
The latter differs by having a lower number of teeth in one transverse row. All other representatives of the genus differ from G. ushunensis by having long salivary glands, a rounded or oval surface of left anterior adductor, a bilaterally symmetrical position of the posterior adductor and by the absence of a velum in the male copulatory organ ( Fig. 6 View FIGURE 6 A–E). G. ushunensis is also the only species among acroloxids from Lake Baikal with scales on the dorsal jaw part.
Distribution and habitat. Gerstfeldtiancylus ushunensis is only known from the type locality and it coexists with G. benedictiae and G. k o t y e n s i s. The new species is most abundant (up to 114 specimens /m2 and 3.81 biomass g/m2, respectively) at the depth of 20–22 m on a bottom composed of multi-layered boulders.

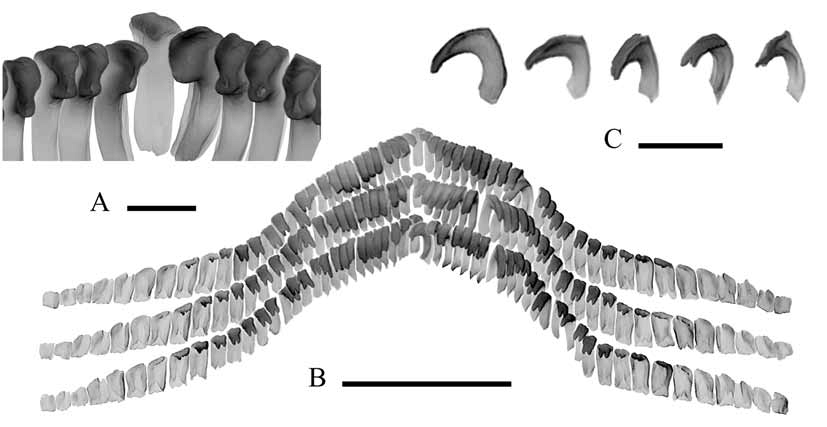
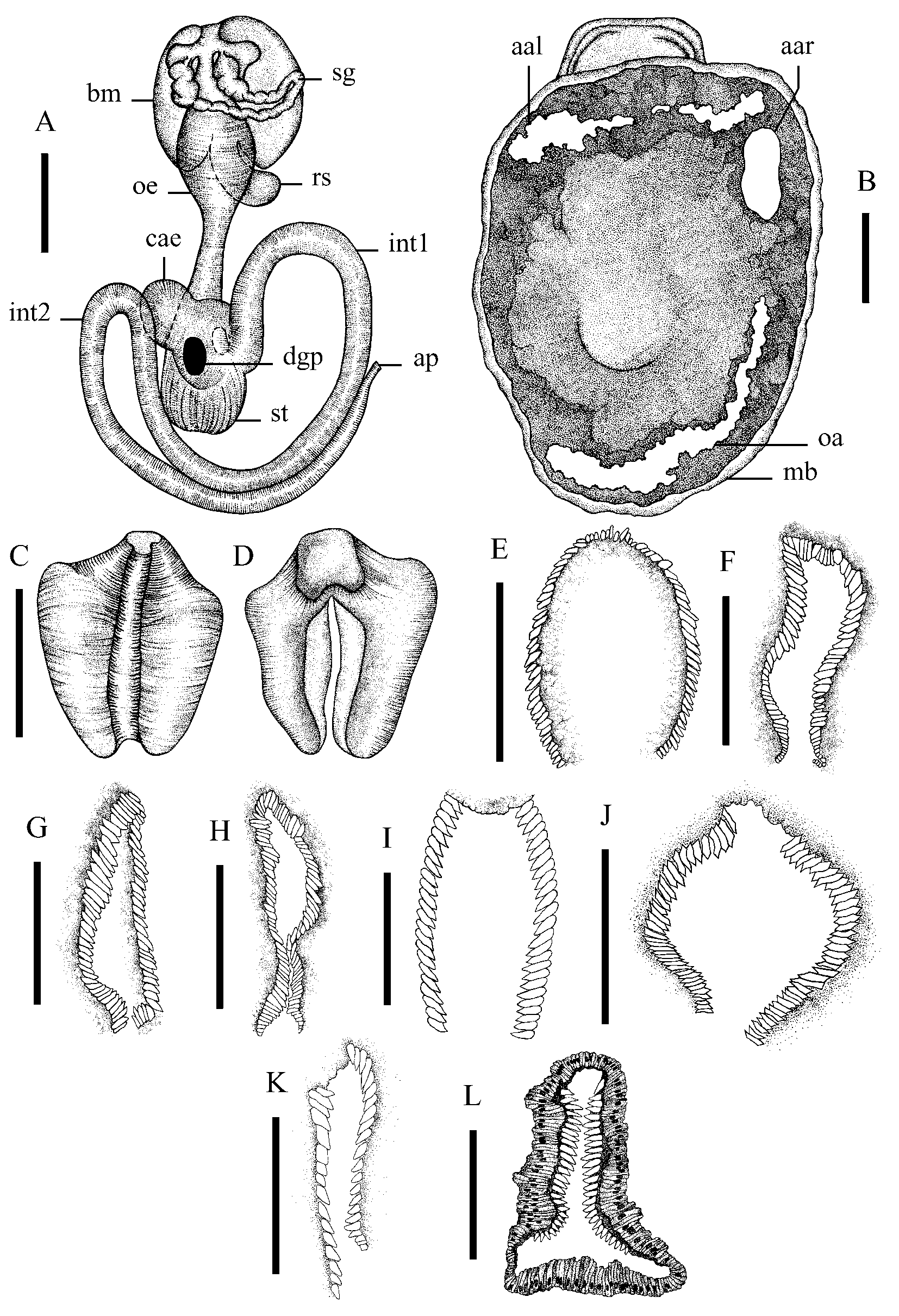
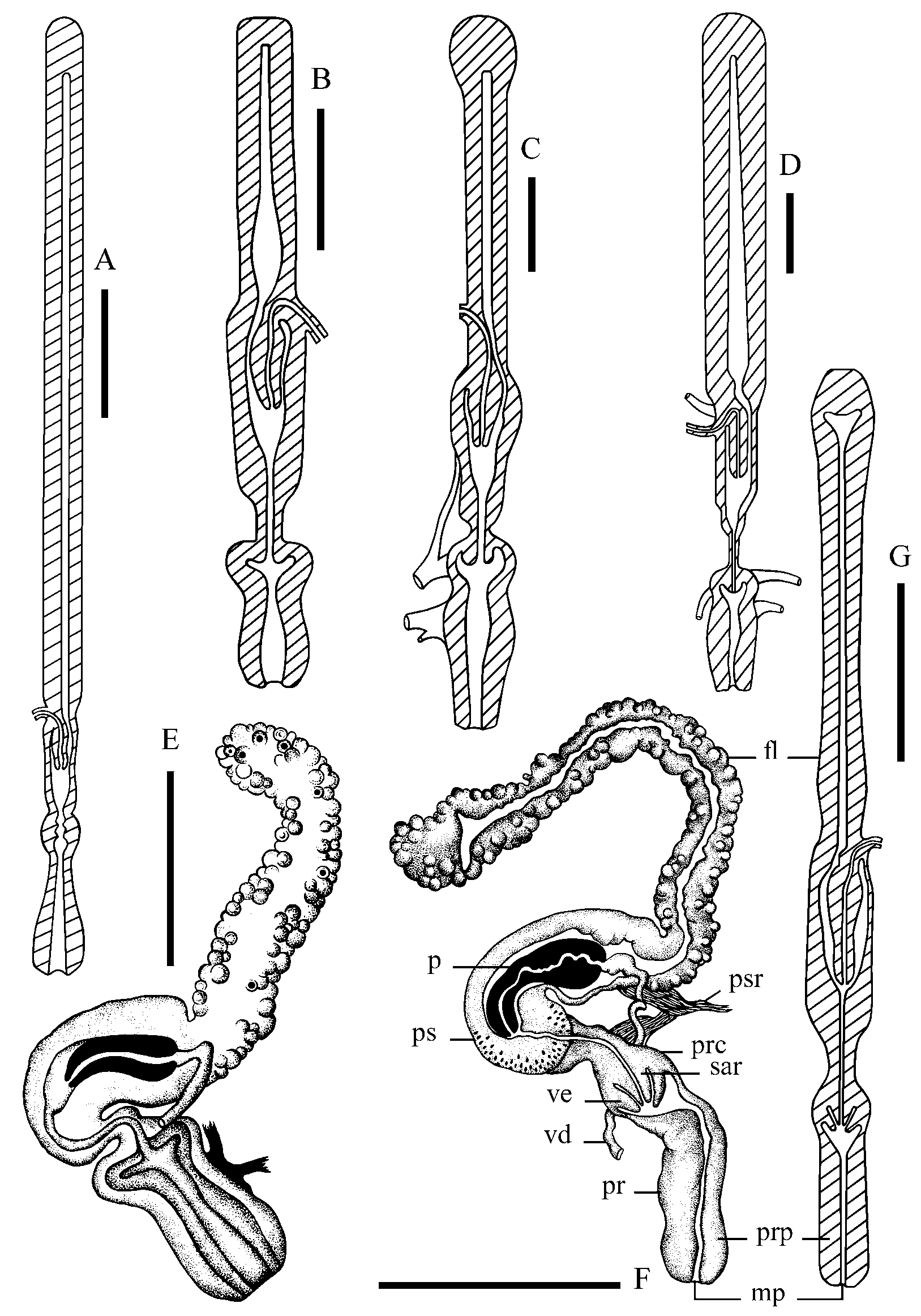
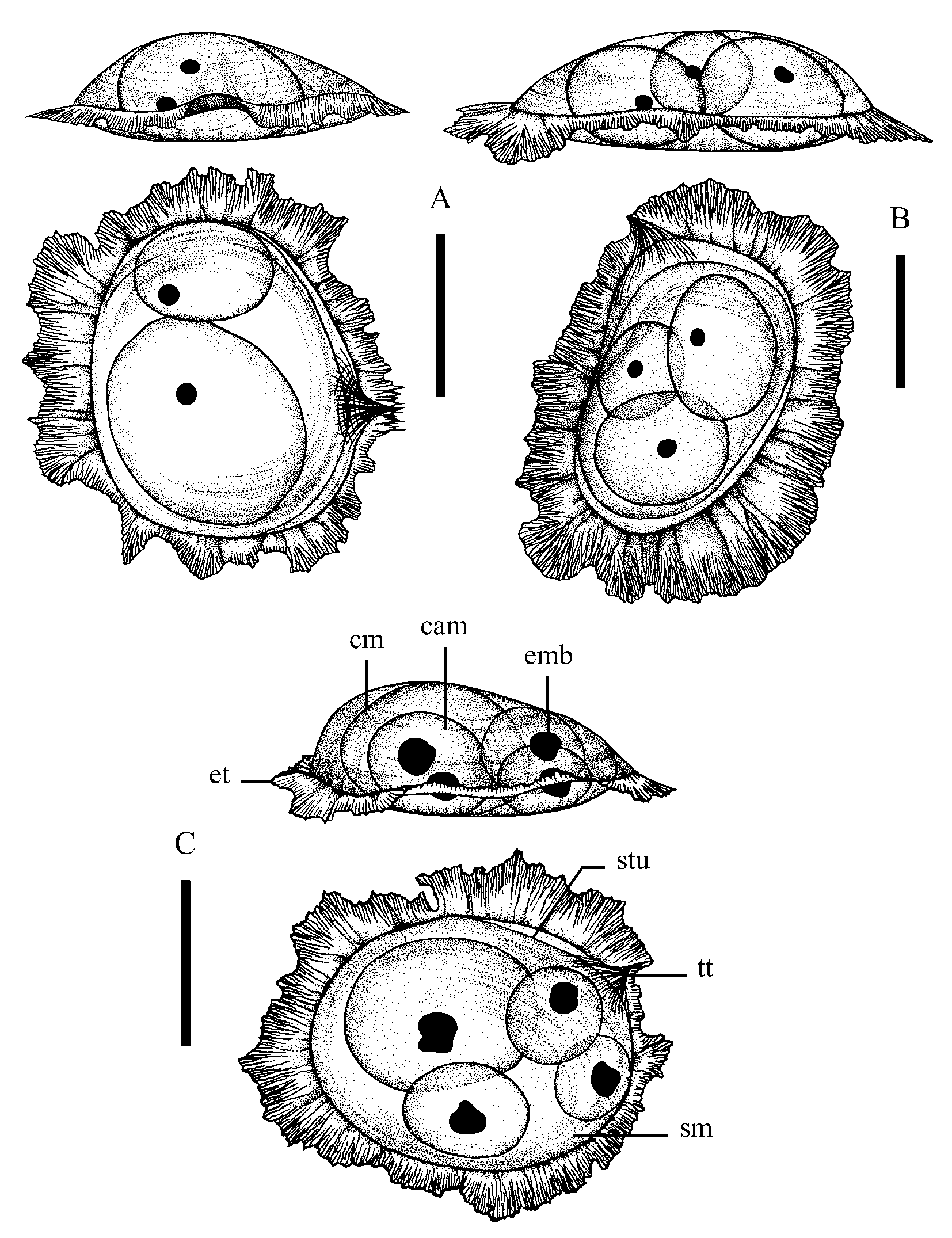
TABLE 1. Shell dimensions of Gerstfeldtiancylus ushunensis, sp. nov. holotype (ZIN RAS 1 / 521 – 2006) and paratypes (in mm). For abbreviations of dimensions see Fig. 2.
| Specimen number | L | La | W | wL | a | H |
|---|---|---|---|---|---|---|
| ZIN RAS 1/521–2006 | 7.20 | 4.20 | 5.80 | 2.50 | 5.50 | 3.00 |
| ZIN RAS 2/521–2006 | 7.40 | 5.00 | 5.90 | 2.90 | 5.70 | 3.15 |
| ZIN RAS 2/521–2006 | 6.95 | 4.55 | 5.55 | 2.50 | 4.95 | 2.85 |
| ZIN RAS 2/521–2006 | 7.20 | 4.60 | 5.70 | 2.70 | 5.30 | 2.65 |
| ZIN RAS 2/521–2006 | 6.90 | 4.35 | 5.60 | 2.70 | 4.75 | 2.55 |
| ZIN RAS 2/521–2006 | 6.40 | 3.85 | 5.10 | 2.50 | 4.40 | 2.20 |
| ZIN RAS 2/521–2006 | 6.05 | 3.60 | 5.10 | 2.50 | 4.40 | 2.10 |
| ZIN RAS 3/521–2006 | 4.00 | 2.45 | 3.30 | 1.65 | 2.90 | 1.20 |
| ZIN RAS 2/521–2006 | 4.70 | 3.05 | 4.10 | 1.90 | 3.50 | 1.80 |
| ZIN RAS 2/521–2006 | 4.50 | 2.85 | 3.90 | 1.70 | 3.40 | 1.85 |
| ZIN RAS 2/521–2006 | 4.50 | 2.90 | 3.80 | 1.80 | 3.30 | 1.50 |
| LIN SB RAS 904 | 2.70 | 1.75 | 2.15 | 0.90 | 1.85 | 0.75 |
| LIN SB RAS 904 | 2.40 | 1.65 | 2.05 | 0.90 | 1.75 | 0.80 |
| LIN SB RAS 905 | 2.95 | 1.85 | 2.35 | 1.10 | 1.90 | 0.75 |
| LIN SB RAS 905 | 3.25 | 2.00 | 2.50 | 1.00 | 2.05 | 0.80 |
| LIN SB RAS 905 | 2.85 | 1.65 | 2.30 | 0.95 | 2.00 | 0.60 |
TABLE 2. Dimensional characteristics of Gerstfeldtiancylus ushunensis, sp. nov. egg masses (in mm).
| No. | Length of syncapsule | Width of syn- capsule | Convexity of syncapsule | Number of egg capsulae | Length of egg capsulae | Width of egg capsulae |
|---|---|---|---|---|---|---|
| 1 | 5.12 | 4.60 | 1.38 | 2 | 1.75–2.79 | 1.25–2.27 |
| 2 3 | 5.26 6.59 | 4.52 4.83 | 1.63 1.86 | 4 3 | 1.54±0.59 (1.06–2.37) 1.96±0.33 (1.59–2.22) | 1.25±0.41 (0.88–1.84) 1.54±0.27 (1.23–1.73) |
| 4 | 5.30 | 4.00 | 1.60 | 5 | 2.25±0.42 (1.57–2.64) | 1.64±0.19 (1.51–1.97) |
| 5 6 | 6.90 7.00 | 5.20 6.50 | 2.30 2.80 | 5 5 | 2.67±0.35 (2.17–2.99) 2.80±0.56 (2.11–3.26) | 1.97±0.16 (1.84–2.18) 2.28±0.48 (1.63–2.79) |
| 7 | 6.00 | 5.30 | 2.94 | 4 | 2.45±0.47 (1.88–2.92) | 1.96±0.59 (1.35–2.77) |
| 8 9 | 5.90 5.30 | 4.20 5.00 | 2.00 1.90 | 3 2 | 3.01±0.1 (2.95–3.13) 2.34–3.27 | 2.11±0.29 (1.81–2.39) 1.65–2.95 |
| 10 | 5.70 | 5.50 | 1.50 | 2 | 2.68–3.49 | 2.25–2.30 |
| 11 12 | 5.70 4.80 | 5.55 4.35 | 2.30 1.40 | 3 2 | 2.74±1.20 (1.97–4.12) 2.60–2.66 | 2.08±0.59 (1.65–2.75) 1.74–2.14 |
| 13 | 5.50 | 4.00 | 2.10 | 3 | 3.15±0.22 (2.92–3.36) | 2.11±0.09 (2.05–2.21) |
| 14 | 5.80 | 5.50 | 2.30 | 3 | 3.06±0.40 (2.61–3.37) | 2.27±0.16 (2.11–2.42) |
| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Gerstfeldtiancylus |