
Fejfarictis valecensis, Bonis & Ekrt & Kunstmüllerová & Martínek & Rapprich & Wagner, 2024
|
publication ID |
https://doi.org/10.5252/geodiversitas2024v46a1 |
|
publication LSID |
urn:lsid:zoobank.org:pub:6982E582-5FCE-4F7A-93A1-43FDB69E59D1 |
|
DOI |
https://doi.org/10.5281/zenodo.10528315 |
|
persistent identifier |
https://treatment.plazi.org/id/4937E6B9-4562-4451-BE1B-94F0CF483EED |
|
taxon LSID |
lsid:zoobank.org:act:4937E6B9-4562-4451-BE1B-94F0CF483EED |
|
treatment provided by |
Plazi |
|
scientific name |
Fejfarictis valecensis |
| status |
n. gen., n. sp. |
Fejfarictis valecensis n. gen., n. sp.
( Figs 3 View FIG , 4 View FIG )
urn:lsid:zoobank.org:act:4937E6B9-4562-4451-BE1B-94F0CF483EED
HOLOTYPE. — Fragmentary left hemi-mandible with p4-m2, alveoli of p1-p3 and partly preserved alveolus of c inf. ( NM-Pv 12300 ; Figs 3 View FIG , 4 View FIG ) .
REPOSITORY. — Department of Palaeontology, National Museum of the Czech Republic, Prague.
ETYMOLOGY. — According to the type locality.
TYPE LOCALITY AND HORIZON. — Valeč-Šibeniční Vrch (= Waltsch-Galgenberg), Northern Bohemia, Czech Republic; early Oligocene (Rupelian), probably MP 21.
DIAGNOSIS. — As for genus.
DESCRIPTION
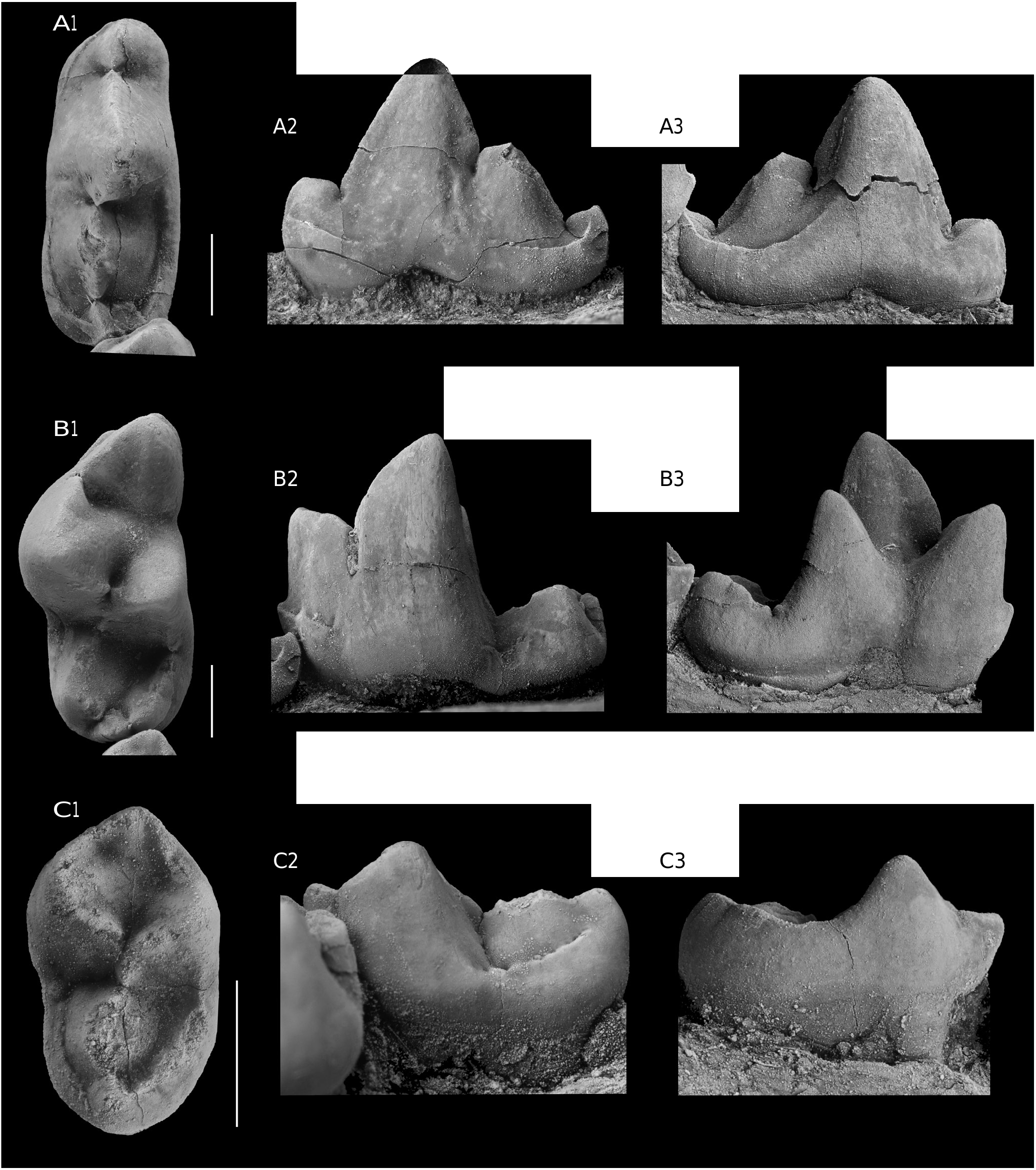
The holotype is the only available specimen. It is a horizontal ramus of a left hemi-mandible ( Fig. 3 View FIG ), missing both a small anterior-most part and almost the entire vertical ramus. p4-m2 (see Table 1 View TABLE for measurements), alveoli of p1-p3 and part of the canine alveolus are preserved. The m3 was not developed. The length of the preserved part of the mandible (measured from anterior-most to distal-most point) is 68.9 mm, the height of the mandible body under m1 (measured on the medial side) is 12.2 mm, the length of p4-m2 (on alveoli) is 20.5 mm, the length of p1-p3 (on alveoli) is c. 17.1 mm (the alveolar margin in front of p4 is slightly damaged).
The first premolar was one-rooted and p2 and p3 had two roots ( Fig. 3B View FIG ). The p4 ( Fig. 4A View FIG ) is elongate relative to m1 (Lp4/Lm1 = 0.89). Its main cuspid is as high as the paraconid of m1, and relatively trenchant; there are mesial and distal accessory cuspids, a well-developed aacd and a far larger pacd; a small cingulid runs along the buccal side of aacd and another one, larger, turns back the pacd as a crest limiting a narrow valley on the lingual side and forming a small cuspid distal to pacd ( Fig. 4A View FIG 3 View FIG ). The crown is asymmetrical, the part distal to the protoconid apex being longer than the proximal one.
The trigonid of the carnassial ( Fig. 4B View FIG ) is high with a quite great height difference between paraconid and protoconid. The former has a cingulid at the buccal base, which gives rise to a narrow trench along the paraconid, interrupted by a thin edge. Lingually, the base of the paraconid contacts the base of the metaconid. The lingual face of the protoconid is marked by a subvertical, central bulging, running from the top to the base, and framed, mesially and distally, by narrow flat surfaces. The metaconid is as high as the paraconid and is relatively distally shifted. As a result, a small part of its posterior portion is visible in a buccal view of the crown. There is a narrowing at the limit between trigonid and talonid, with a large notch between the distal face of the protoconid and the hypoconid. The latter, preceded by a small cristid obliqua, is elongate; its lingual face is flat. It seems to be slightly affected by abrasion, but it is hard to reconstruct the original surface and shape with certainty. Currently, there is no convexity, which is seen in the lingual face of the protoconid. Preceded by a small cusplet followed by a short ridge at the disto-lingual base of the metaconid, the entoconid is also relatively high. The hypocristid and entocristid both encircle an elongate deep talonid basin, which is open distally, despite the presence of a hypoconulid separated from the hypocristid by a deep notch.
The m2 ( Fig. 4C View FIG ) is relatively short (Lm2/Lm1 = 0.49). The trigonid is complete and clearly higher than the talonid. The protoconid is slightly similar to that of m1, with a convex buccal face and a lingual one more flattened with a central vertical pillar (less developed than in m1); the preprotocristid joins a modest paraconid by the preparacristid.The metaconid has a thick postmetacristid which is separated by a notch from the postprotocristid; a short and thick cristid obliqua joins a globular dome-shaped hypoconid of which the top is missing. Like in m1, a weak ridge precedes the entoconid, a thick entocristid, which turns the talonid to join the hypoconulid, closing the oval mesio-distally elongate talonid basin. In fact, the entoconid appears like a thickening of the cristid and the hypoconulid is also only marked by a thickening of the cristid.
That dentition, by the thick cristids, looks like that of a mesocarnivore, with cutting (especially in m1) and grinding functions, the latter being very-well marked in m2.
COMPARISONS
Fejfarictis valecensis n. gen., n. sp. was carnivorous, judging by the morphology of its dentition. Carnivores were present in Europe during the Eocene and the beginning of Oligocene, with the Hyaenodonta and the Carnivoramorpha . The former are very different from the Valeč hemimandible in the structure of the lower molars, whose size increases from m1 to m3. The Valeč hemimandible belongs to the latter, and more precisely to the Carnivoraformes and the clade Carnivora (see Solé et al. 2014, 2022). Viverravidae , present in Europe in early Eocene, differ from Carnivoraformes (and therefore also from Fejfarictis n. gen.), among others, in the morphology of m2 with elongated talonid and enlarged hypoconulid ( Flynn et al. 2010: 37).
We know the Carnivoraformes in Europe from the latest Paleocene ( Solé et al. 2016). In the late Eocene (MP 16), they are present with Simamphicyon Viret, 1942 , Quercygale Kretzoi, 1945 , Paramiacis Matthis, 1985 .
The type and only species of Simamphicyon , S. helveticus (Pictet & Humbert, 1869) is far larger than the Valeč form; it was sometimes considered to be an amphicyonid (see Springhorn 1977), but recently, it was placed into basal carnivoraforms ( Tomiya & Tseng 2016; Solé et al. 2022). It differs from Fejfarictis n. gen. also by the relatively less elongate p4 and the quite trenchant talonid of m1.
The larger species of Paramiacis , P. teilhardi Matthis, 1987 , is smaller than F. valecensis n. gen., n. sp. Its maximum m1 length can reach only 5.1, while it is 9.1 for F. valecensis n. gen., n. sp. The p4 is relatively shorter, high and pointed, without accessory cuspids. The trigonid of m1 is relatively higher and with more pointed cuspids; the m2 is relatively larger and the paraconid seems to be very small or absent. Paramiacis has an m3.
Quercygale angustidens ( Filhol, 1872) was recorded in several late Eocene localities. The revision of its phyletic position places the taxon as “the most derived miacid” ( Wesley-Hunt & Werdelin 2005), just before the crown-group of Carnivora in a cladogram, after a study of the cranial characters. A similar result occurs in an analysis of the Carnivoraformes, using 246 characters ( Solé et al. 2014). The m3 of Quercygale is lost, as is that of Fejfarictis n. gen. Nevertheless, Quercygale differs from our fossil in the shape of the anterior part of the mandible, with a marked flange and a large mental foramen, p4 without anterior accessory cuspid, shorter talonid of m1 and smaller m2.
In Europe, the end of Eocene is marked by the arrival of a small amphicyonid, Cynodictis Bravard & Pomel, 1850 . The type-species, C. lacustris Bravard & Pomel, 1850 , comes from the locality La Débruge (MP 18; Bravard & Pomel 1850), but the genus was also recorded in later Eocene localities (MP 19; Aguilar et al. 1997; Le Verger et al. 2020). Like all Eocene and Oligocene amphicyonids, Cynodictis has a lower m3, although some specimens may have lost m3 ( Teilhard de Chardin 1915), but that character must have been rare. In the locality Sainte-Néboule de Béduers, the only locality where it is possible to observe a Cynodictis population (minimum number of individuals = 24), there is no individual without m3 ( Bonis 1978). The size of the Valeč specimen is compatible with that of the Cynodictis of Sainte-Néboule de Béduer. Nevertheless, the proportions are different for p4; the index [(Lp4/Lm1) × 100] is 89 for Fejfarictis n. gen. and varies from 44 to 69 in Cynodictis , while for m2 the same index is 49 for Fejfarictis n. gen. and varies from 44 to 69 in Cynodictis , placing the Valeč fossil among the smaller indices of Cynodictis . In Cynodictis , there is no well-developed anterior accessory cuspid on the p4, only sometimes a small cingulid spur, and the distal accessory cuspid is lower and smaller.
Another amphicyonid, Storchictis Bonis, 2020 , differs from Fejfarictis n. gen. in the same characters as does Cynodictis ( Bonis 2020) . This taxon comes from the old collections of the “Phosphorites du Quercy”, and its precise stratigraphic level is unknown, but it could be late Eocene or early Oligocene.
The“Grande Coupure”( Stehlin1909) is a milestone separating in time, in Western Europe, the Eocene terrestrial mammalian faunas from the Oligocene ones. Many genera of carnivorans, Caniformia and Feliformia were present among the new migrants ( Solé et al. 2022 and references therein). In the Caniformia, the Amphicynodontidae were the most abundant. Their morphology does not correspond to that of Valeč specimen. The teeth are generally lower with a more developed grinding surface, such as Amphicynodon Filhol, 1881 and especially Pachycynodon Schlosser, 1888 . All have the m3 preserved. The musteloids ( Mustelictis Lange, 1969 ) have lost m3, but the carnassial is relatively lower with a lesser height difference between paraconid and metaconid. Taxa most similar to Fejfarictis n. gen. belong to the aeluroids, in which m3 is always lacking. Most of these genera were described by Filhol (1872a, b, 1876 -1877, 1880), using material from the “Phosphorites du Quercy”.
Several aeluroid genera are known in the Western European early Oligocene (revision by Hunt 1998). The most common is Stenoplesictis Filhol, 1880 , with three species: S. cayluxi Filhol, 1880 , S. minor Filhol, 1882 and S. crocheti Peigné & Bonis, 1999 . Stenoplesictis differs from Fejfarictis n. gen. in having the p4 shorter and higher than m1, with a less developed distal cingulid; the m1 with a more reduced metaconid, with a lesser height difference between paraconid and protoconid, and a shorter talonid; m2 is shorter with thinner cristids, more pointed cuspids and generally single-rooted. The mandible of Palaeoprionodon Filhol, 1880 is smaller and thinner than that of Stenoplesictis , but its overall morphology is quite similar. Haplogale Schlosser, 1888 is distinguished by a low and elongated mandibular corpus; its m1 is high with a very reduced metaconid, and clearly different from that of Valeč specimen. Stenogale Schlosser, 1887 has also a shorter p4, a m1 with a very reduced metaconid and a smaller and more trenchant talonid, and a small one-rooted m2. We may write almost the same for Viretictis Bonis, Peigné & Hugueney, 1999 . Especially its m1, with a more reduced metaconid and a trenchant hypoconid, differs clearly from that of Fejfarictis n. gen.
These aeluroids, most of which come from the old collections of the phosphorites of Quercy ( Teilhard de Chardin 1915), have no precise reference levels, but some were found with new excavations dated to the Oligocene. As underlined by Hunt (1998: 7) “An interesting aspect of the Quercy aeluroid fauna is the tendency of most taxa to develop a hypercarnivorous dentition”. This tendency does not appear in the Valeč carnivoran.
Another Quercy carnivoran may be compared with Valeč aeluroid. It is Anictis Kretzoi, 1945 , figured by Schlosser (1887) and Hunt (1998). Anictis differs from the other aeluroids of the Quercy. It is less derived and certainly closer to the stem group of the Feliformia . It is known through two hemi-mandibles figured by Schlosser (1887: pl. IX, fig. 54) and Hunt (1998: fig. 3), respectively. Both come from the old collections without precise dating, although Anictis would have been recorded as early Oligocene in age (MP 21; Lange-Badré 2006). Anictis is relatively primitive when compared to the other Quercy aeluroids, and forHunt (1998: 7), it has a “moderately plesiomorphic dentition”. It differs from Fejfarictis n. gen. by these characters: p4 is simple, pointed, no mesial accessory cuspid and smaller distal one, without distal developed cingulid, the height difference between paraconid and protoconid of m1 is similar to that of Valeč, but there is no hypoconulid on m1; m2 is two-rooted but has “the three trigonid cusps positioned at the points of an equilateral triangle” ( Hunt 1998) and the protoconid is clearly higher.
Other aeluroids, from the late Eocene or early Oligocene of Asia, may be compared with the Valeč carnivoran. An American Museum of Natural History expedition in Mongolia enabled Matthew & Granger (1924) to publish several new Aeluroidea taxa and the ubiquitous genus Palaeogale Meyer, 1846 ( Matthew & Granger 1924). The aeluroids are “ Cynodictis ” elegans Matthew & Granger, 1924 for a mandible with the canine and the premolars, Palaeoprionodon gracilis Matthew & Granger, 1924 for a fragment of mandible with p3, partially broken off m1 and Viverravus constans Matthew & Granger, 1924 for a piece of mandible with m1-m2.
Because only premolars are known for “ Cynodictis ” elegans , the possibility of direct comparison with F. valecensis n. gen., n. sp. is limited. Based on the given drawings ( Matthew & Granger 1924: fig. 6A; Hunt 1998: fig. 24A), anterior accessory cuspid of p4 is much less developed than in F. valecensis n. gen., n. sp. and “ C. ” elegans p4 seems to lack a lingual cingulid. Matthew & Granger (1924) as well as Hunt (1998) suppose that this species could be conspecific with “ P. ” gracilis (see below), but they leave the question open.
Other material of stenoplesictoid and Palaeogalidae from Mongolia was described by Huang (1993). Fragments of mandibles named Cynodictis elegans Matthew and Granger, 1924 has a p4 without aacd and two other fragments, figured as Palaeoprionodon , bear a p4 shorter (relative to m1) than that of the Valeč specimen, and an m1 with a smaller metaconid. Both teeth have a swollen buccal cingulid. These taxa are different from the Fejfarictis n. gen.
Asiavorator Spassov & Lange-Badré, 1995 is known with A. altidens Spassov & Lange-Badré, 1995 as a type species, from the Mongolian Oligocene. The species has a simple, shorter p4 without mesial accessory cuspid and a smaller distal accessory cuspid; the m1 is more trenchant with a large paraconid-protoconid, a reduced metaconid and a cutting, non-basined talonid ( Spassov & Lange-Badré 1995: figs 1-3). Hunt (1998) assigned Palaeoprionodon gracilis to this genus and even synonymised both species under the name Asiavorator gracilis . Asiavorator is more specialized towards hypercarnivory than Fejfarictis n. gen.
Dashzeveg (1996) published other Mongolian fossils, two hemi-mandibles, under the generic name Stenoplesictis , S. indigenus Dashzeveg, 1996 (now Alagtsavbaatar Egi, Tsubamoto, Saneyoshi, Tsogtbaatar, Watabe, Mainbayar, Chinzorig & Khatanbaatar, 2016 ) and S. simplex Dashzeveg, 1996 from the late Eocene and/or the early Oligocene respectively; they have a morphology seemingly close to that of F. valecensis n. gen., n. sp. ( Dashzeveg 1996: figs 2-3). Both species have an elongate p4 with well-marked mesial and distal accessory cuspids, a cingulid turning back the crown, an overall profile of m1 similar to that of Valeč m1 and a double-rooted m2. Nevertheless, A. indigenus differs from F. valecensis n. gen., n. sp. by the lower entoconid of m1, absence of hypoconulid and trenchant talonid of m2 (for the latter character see Egi et al. 2016). “ Stenoplesictis ” simplex Dashzeveg, 1996 was known by a right hemi-mandible with p3-m2; it differs from F.valecensis n. gen., n. sp. by the shorter talonid of p4, the thick cingulid along the buccal face of m1, the more reduced metaconid of m1, the three conic cuspids of the m2 trigonid and its narrow talonid. Both species have a p4 more symmetrical than that of Valeč p4.
Hunt (1998) described a new genus Shandgolictis Hunt, 1998 with Viverravus constans as its type species. Shandgolictis differs from Fejfarictis n. gen. by a low, short, reduced and not basined talonid of m1 ( Hunt 1998), which has also a thick cingulid all along the buccal face; m2, although bi-rooted, is smaller with a clear tricuspid trigonid and a short and narrow talonid tapering distally. He brings “ Stenoplesictis ” simplex and “ S. ” indigenus close to Shandgolictis constans rather than to Stenoplesictis (see also Peigné & Bonis 1999). This view was followed by Morlo & Nagel (2007) who even used the binomen Shandgolictis simplex .
Egi et al. (2016), in the newest revision of the early feliformians from Mongolia, describe new material and review the preceding taxa. They consider, following Hunt (1998), the mandible fragment figured by Matthew & Granger (1924: fig. 6E) as Palaeoprionodon gracilis as belonging to Asiavorator (but contra Hunt (1998), with “ Stenoplesictis ” simplex as its junior synonym). “ S. ” indigenus is designated as type species of a new genus, Alagtsavbaatar . The differences between this new genus and Fejfarictis n. gen. were given above for A. indigenus .

| MP |
Mohonk Preserve, Inc. |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Feliformia |
|
InfraOrder |
Aeluroidea |
|
Family |
|
|
Genus |