
Escharoides jacksoni ( Waters, 1900 )
|
publication ID |
https://doi.org/ 10.1080/00222930601162878 |
|
persistent identifier |
https://treatment.plazi.org/id/03C23512-D11B-4E70-E1F9-C193D4A00446 |
|
treatment provided by |
Felipe |
|
scientific name |
Escharoides jacksoni ( Waters, 1900 ) |
| status |
|
Escharoides jacksoni ( Waters, 1900) View in CoL
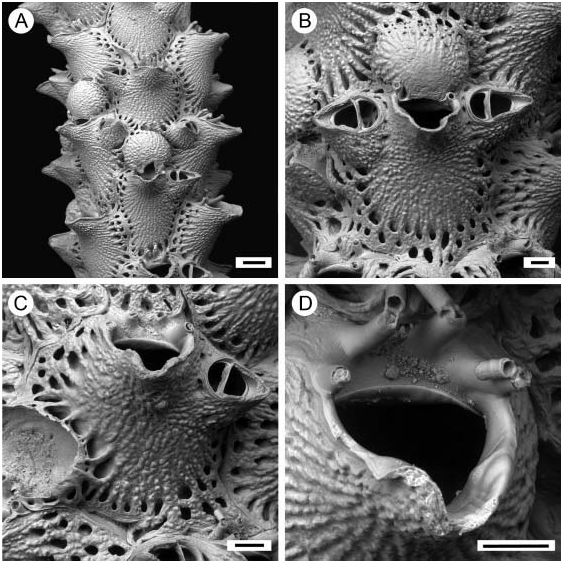
( Figures 3 View Figure 3 , 4 View Figure 4 )
Smittina jacksoni Waters 1900, p 87 , Plate 12, Figure 18.
Mucronella coccinea ( Abildgaard, 1806) : Bidenkap 1897, p 624, Plate 25, Figures 5 View Figure 5 , 6.
Escharoides jacksoni var. rostrata Kluge 1946, p 201 , Figure 6; Kluge 1962, p 569, Figure 402; Kluge 1975, p 693, Figure 402.
Escharoides jacksoni: Kluge 1962, p 568 View in CoL , Figure 401; Kluge 1975, p 692, Figure 401; Powell 1968, p 2294, Plate 5, Figure B; Bille-Hansen 1962, p 33.
Type material examined
Holotype: MM 4307 , 77 ° 559N, 55 ° 109E, collected during Jackson-Harmsworth Expedition, 7 October 1897, Waters Collection.
Other material examined
NHM 2006.07.31.2, Laptev Sea, 74 ° 30.09N, 137 ° 05.09E, Transdrift 1 project, Stn 48, 18 August 1993, 22 m depth, dredge, coll. Schmid GoogleMaps ; ZI 30/2945, East Siberian Sea, Stn 25, 23 August 1937, ‘‘ Sadko’ ’, det. Kluge, coll. Gorbunov ; ZI 50/354, NE Polar Basin, Stn 73, 1948, det. Petrovskaya; ZI 7, Fram Straight , Stn 43, 1901, ‘‘ Zaria’ ’, det. Kluge, coll . R.P.E.; ZI 10, Laptev Sea, in front of Pronchischev Bay, Stn 34, ‘‘ Sibiriakov’ ’, trawl, 14 September 1933, det. Kluge, coll. Gorbunov ; ZI 15, Kara Sea, O-b Ruskij– Dickson Bay, Stn 40, ‘‘ Sibiriakov’ ’, trawl, 27 August 1933, det. Kluge, coll. Gorbunov ; ZI 65/50534, Chukchi Sea, depth 39 m, Stn 17, 13 August 2004, ‘‘ Prof. Khromov’ ’, coll. Sirenko, Gagaev, Denisenko Collection ; CMN 2006-0013 About CMN , southwestern Baffin Island , 69 ° 55.79N, 80 ° 199W, Stn 820, depth 73 m, 28 August 1956, Powell Collection ; CMN 2006-0014 About CMN , Cornwallis Island , 72 ° 44.19N, 94 ° 069W, Stn 62-2014, depth 35 m, 25 July 1962, Powell Collection ; CMN 2006-0015 About CMN , Nunavut, Frozen Strait, 65 ° 329N, 84 ° 529W, Stn 61-39, depth 38–42 m, 27 August 1961, Powell Collection ; CMN 2006-0016 About CMN , Creswell Bay , 72 ° 46.59N, 94 ° 15.59W, Stn 62-2052, depth 59–62 m, 11 August 1962, Powell Collection GoogleMaps .
Description
Colony forming a thick unilaminar crust ( Figure 3A View Figure 3 ). Ancestrula ( Figure 3D View Figure 3 ) tatiform, oval, with narrow cryptocyst, broad proximal gymnocyst, and 11 spines.
Autozooids irregular in shape, separated by deep grooves, large, 0.88–1.33 mm long (mean 1.02 mm, n 520) by 0.58–0.88 mm wide (mean 0.69 mm, n 520). Frontal shield finely granular, bordered by a single or double row of small, round areolae. Orifice suborbicular; distal border with a broad, smooth, suboral shelf; proximal edge developed as a thick, projecting peristome, sometimes with one or two thickened lateral denticles around proximal inner margin ( Figures 3B View Figure 3 , 4B View Figure 4 ), and often with a median notch on the rim. Denticles striated when present. Oral spines short, usually numbering four, the outer two fused with the peristome, only two visible in ovicellate zooids. Avicularia typically paired at distolateral corners of autozooids, sometimes single or absent, large, 0.22–0.32 mm long (mean 0.27 mm, n 520) by 0.13–0.20 mm wide (mean 0.15 mm, n 520), directed laterally, rostrum long, tapering distally, rarely hooked at tip; avicularian chamber well developed, pores often present at the base and rarely on chamber itself; mandible elongate-triangular, crossbar calcified.
Ovicell ( Figures 3B View Figure 3 , 4B View Figure 4 ) hyperstomial, globular, finely granular, slightly longer than broad, large, 0.35–0.46 mm long (mean 0.40 mm, n 520) by 0.37–0.44 mm wide (mean 0.41 mm, n 520); pseudopores scattered over distal part, bordered by a single or double series of small, round areolae.
Distribution
An Arctic, circumpolar species recorded from the Greenland, Barents, Kara, Laptev, East Siberian, and Chukchi Seas; and the Canadian Arctic ( Kluge 1975; current account).
Remarks
Bille-Hansen (1962) and Hayward and Ryland (1999) synonymized Escharoides bidenkapi ( Kluge, 1946) with E. jacksoni ( Waters, 1900) . However, their analyses were based on small sample sizes, and the synonymy is not supported by a more detailed comparison. Our broader investigation using SEM permits us to distinguish E. bidenkapi ( Figure 2 View Figure 2 ) and E. jacksoni ( Figures 3 View Figure 3 , 4 View Figure 4 ) as separate species. Both species have a similar peristome shape, but there is large variation both among colonies and within colonies of Escharoides in this character. However, a few skeletal morphological characters do allow us to distinguish these two species with confidence. In general, the peristome of E. bidnekapi is much more complex than that of E. jacksoni . In E. jacksoni the peristome is never produced into three prominent denticles (cf. Figure 2C View Figure 2 ). On average, autozooids of E. bidenkapi are smaller than those of E. jacksoni , as are the avicularia. Also, in the majority of cases, avicularia of E. bidenkapi are directed distolaterally, whereas those of E. jacksoni are typically directed laterally. The frontal shield texture of E. bidenkapi is polygonal, whereas that of E. jacksoni is finely granulated, although in early astogeny E. jacksoni also has frontal shields with a polygonal texture (see Figures 2D View Figure 2 , 3D View Figure 3 ). A very stable character of skeletal morphology among the specimens studied is the structure of the distal edge of the suboral shelf. In E. bidenkapi this is crenulated, whereas in E. jacksoni it is always smooth. Escharoides jacksoni has more bulbous avicularian chambers, which are better demarcated from the autozooidal frontal shield than in E. bidenkapi .
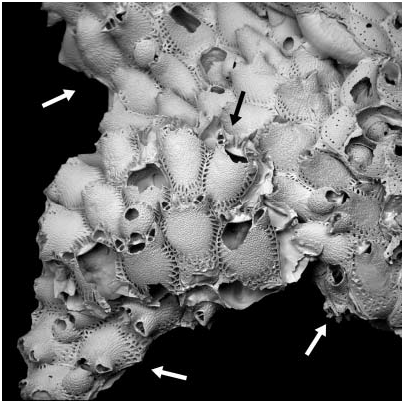
Escharoides bidenkapi and E. jacksoni overlap in geographical distribution but whereas E. jacksoni is circumpolar in distribution, E. bidenkapi does not occur in some parts of the Arctic (e.g. Canadian Arctic, Chukchi Sea). In our study material, we found one instance of the two species growing together on the same substratum ( Figure 5 View Figure 5 ). The adjacent colonies show the distinctive characters mentioned above for the two species, which cannot be ecophenotypic as the colonies are juxtaposed. This specimen therefore supports the assertion that E. bidenkapi and E. jacksoni are two separate species.
According to Kluge (1946), E. jacksoni var. rostrata ( Figure 4 View Figure 4 ) exhibits an atypical peristome shape (‘‘strongly raised forward and narrows toward the end’’). However, the majority of skeletal characters (frontal shield, avicularian size and shape, smooth-edged suboral shelf) allow us to subsume this taxon into E. jacksoni (Waters) .


| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Escharoides jacksoni ( Waters, 1900 )
| Kuklinski, Piotr, Taylor, Paul D. & Denisenko, Nina 2007 |
Escharoides jacksoni:
| Kluge GA 1975: 692 |
| Powell NA 1968: 2294 |
| Kluge GA 1962: 568 |
| Bille-Hansen K 1962: 33 |
Escharoides jacksoni var. rostrata
| Kluge GA 1975: 693 |
| Kluge GA 1962: 569 |
| Kluge GA 1946: 201 |
Smittina jacksoni
| Waters A 1900: 87 |
Mucronella coccinea ( Abildgaard, 1806 )
| Bidenkap O 1897: 624 |