
Errina capensis Hickson, 1912
|
publication ID |
https://doi.org/10.11646/zootaxa.3691.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E98CE6DF-AF3B-4AAA-95CB-8ACD615C9FCC |
|
DOI |
https://doi.org/10.5281/zenodo.5619737 |
|
persistent identifier |
https://treatment.plazi.org/id/955B87C9-A165-DD36-FF22-FF16F0D92D63 |
|
treatment provided by |
Plazi |
|
scientific name |
Errina capensis Hickson, 1912 |
| status |
|
Errina capensis Hickson, 1912 View in CoL
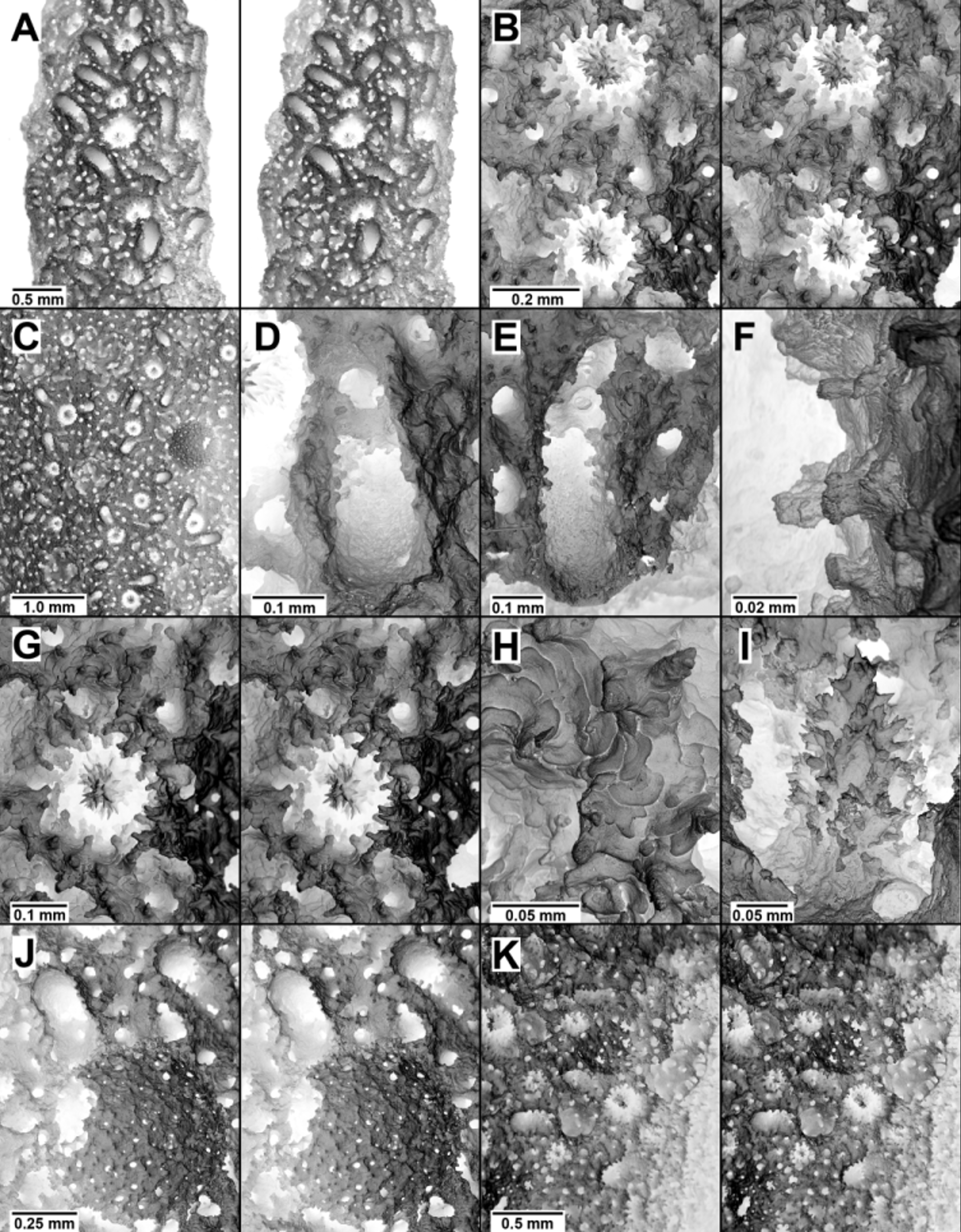
Figs. 1 View FIGURE 1 G, 8A–K, 26
Errina (Labiopora) capensis Hickson, 1912: 886 –887, 890, pl. 95, fig. 7, pl. 96, fig. 15. Errina capensis: Hickson, 1912: 894 (listed).—Broch, 1942: 38, 40 (key).—Boschma, 1957: 52; 1963a: 337 (listed); 1966b: 112.—Cairns, 1983b: 428 (listed).—Williams, 1986: 14 (colour drawing).
Errina (Eu-Errina) capensis: Broch, 1942: 53 –54, text-fig. 16, pl. 4, fig.15 (redescription). Errina (Labiata) capensis: Broch, 1942: 91 .
Types and Type Locality. Three female syntypes, preserved in alcohol, are deposited at the MNHN; one syntype branch (illustrated by Broch, 1942) is also deposited at the Zoological Museum, Oslo (B884); one syntype fragment is at the Manchester Museum; and one syntype fragment is at the BM (1950.1.11.90). Type Locality: off Cape of Good Hope, 30 fm (= 55 m)(see Hickson 1912: 890).
Material Examined. Types; MN SM180, 1, SAM H3154; MN SM185, 1, SAM H3153; MN XX129, 1 male, 1 female, 1 indet. colonies, SAM, and SEM stubs 1607–609, USNM; PF 503, 1, SAM H3090; PF 808, 2, SAM H1221; PF 11555, 1 colony, SAM H1237; PF 12104, 1 female, 3 indet. colonies, SAM H3093; PF 12314, 2 female branches, SAM H1219; PF 13394, 24, SAM H1233; PF 13465, 1 colony, SAM H1234; PF 13476, 19 male branches, SAM H1228 and, and 3 female branches and SEM stub 1666, USNM 76521 and 76522, and four branches, BM1980.9.19.12; PF 13599, 4 colonies, SAM H1232; PF 13959, 9 colonies, SAM H1224; PF 15614, 1, SAM H3092; Valdivia 100, 1, ZMB; off Cape of Good Hope, Manchester Museum.
Description. Colonies are uniplanar, having equal dichotomous branching, the axils between branches being fairly narrow; there is no branch anastomosis and no symbiosis with commensal polychaetes. The largest specimen ( MN XX129) is 8.2 cm in height and 10.5 mm in basal stem diameter. Distal branches are blunt and slightly flattened to elliptical in cross section. The coenosteal texture is basically radial-imbricate, consisting of broad smooth platelets ( Fig. 8 View FIGURE 8 H), but the coenosteal strips are narrow and poorly defined, bordered by rather large coenosteal pores (up to 58 µm in diameter), which confer a somewhat porous aspect to the branch surface. Furthermore, the coenosteum bears numerous small (up to 78 µm), sharp spines ( Fig. 8 View FIGURE 8 G) that project perpendicular to the branch surface, giving it a spiny texture. Colonies are light orange in colour, the branch tips, cores, and ampullae usually being white.
Gastropores are uniformly distributed on all branch surfaces and are circular in shape, ranging from 0.20–0.30 mm in diameter; there are no bordering gastropore lips. The gastrostyles are stout and bullet-shaped ( Fig. 8 View FIGURE 8 I), up to 0.24 mm in height, with a L:D of 1.6–2.0, and occupy the entire gastropore tube, the distal tip sometimes even rising slightly above the coenosteal surface ( Figs. 8 View FIGURE 8 B, G). The gastrostyle is covered with tall (up to 34 µm) pointed spines, which are often fused into longitudinal or oblique ridges. The inner face of the upper half of the gastropore tube bears a wide diffuse ring palisade composed of blunt elements up to 44 µm in height. These elements continue to the uppermost part of the gastropore tube and are almost indistinguishable from the coenosteal spines ( Figs 8 View FIGURE 8 B, G). The horseshoe-shaped dactylopore spines ( Figs. 8 View FIGURE 8 D, E) are quite abundant, covering all surfaces of the branch, but better developed on one side (by definition, the anterior) and lateral branch edges. Near the branch tips the dactylopore spines are uniformly adcauline in orientation and discrete, but farther away from the tip the orientation of their dactylotomes may be lateral or oblique, and several dactylopore spines may be fused laterally into short rows. The dactylopore spines are fairly short (only 0.14–0.29 mm in height), and thin-walled (30–50 µm), about 0.20–0.25 mm in total width, and 0.4–0.5 mm long; the outer wall of the dactylopore spines are often buttressed with several thin ridges, which give the upper spine a serrate edge. Internally, each dactylopore spine has a single, thin, flat platform on its internal upper edge ( Fig. 8 View FIGURE 8 D, E), which blocks about half of the dactylotome. Furthermore, each dactylopore spine has two lateral (but no medial) dactylostyles, the elements being 15–24 µm in height ( Fig. 8 View FIGURE 8 E).
The female ampullae are prominent, hemispherical structures 0.6–1.0 mm in diameter, when mature having a circular lateral efferent pore about 0.30 mm in diameter ( Fig. 8 View FIGURE 8 J). They occur on the anterior and lateral branch faces, sometimes in great abundance. Male ampullae are also superficial but smaller ( 0.45–0.50 mm in diameter), having a porous structure and small apical efferent pores ( Fig. 8 View FIGURE 8 K). They concentrate on the anterior surface, often in clusters.
Comparisons. Among the 27 Recent species of Errina (Appeltans, et al. 2012; Cairns 1999), only two others are known from the Indian Ocean. Errina capensis is similar to the Subantarctic E. kerguelensis Broch, 1942 in gross colony shape and colour, but differs in almost all other characters (see Cairns 1983a). The other species, E. aspera mascarina Boschma, 1965 , from Mauritius, differs in having a white corallum, much taller dactylopore spines, a longer gastropore tube and taller gastrostyles, although the styles do not reach the branch surface, and in lacking a ring palisade and coenosteal spines. Boschma (1965) gave convincing evidence that this subspecies ( Errina aspera mascarina ) was different from the Mediterranean nominal subspecies, although Zibrowius & Cairns (1992: 48) suggested that this specimen was collected in the Mediterranean, not off Mauritius.
Remarks. Despite several references to this species and a host of combinations (see synonymy), until now this species was known only from the type material. This is the first treatment that employs SEM, including the first description of the ampullae, as well as adding several new distributional records. Although no commensal polychaetes live with this species, it does commonly harbor attached barnacles.
Distribution. Known from coastal waters of South Africa from off the Cape of Good Hope to just north of Durban ( Natal Province) (Fig. 26), 40– 174 m.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |