
Eoatrichops jeanbernardi, Myskowiak, Justine & Nel, André, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3869.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:EE009703-DC4D-44C5-9A98-C27DB30B0556 |
|
DOI |
https://doi.org/10.5281/zenodo.6138979 |
|
persistent identifier |
https://treatment.plazi.org/id/6343CA23-6A18-BF76-FF5B-FC61F5950FDF |
|
treatment provided by |
Plazi |
|
scientific name |
Eoatrichops jeanbernardi |
| status |
sp. nov. |
Eoatrichops jeanbernardi View in CoL sp. n.
( Figs 1–13 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 )
Material. Allotype female specimen MNHN.F.A51152 (PA 106 1/2) (nearly complete with apices of some legs missing, together with a badly preserved adult Blattodea, fossilised at the surface of the amber piece). Holotype male specimen MNHN.F.A51153 (PA 9055 1/6) (nearly complete, with wings partly missing, together with three adult Chironomidae , and two incomplete female specimens, both paratypes of the same species, numbers PA 9055 2/6 and PA 9055 3/6). Material stored at the Laboratory of Paleontology, MNHN, Paris, France. The female allotype is partly destroyed with all legs cut at tibia or tarsi level, except one posterior leg, and ventral part of head broken. The male holotype has its wings partly destroyed, but with venation of its basal half still visible. The female paratype PA 90552/6 is also partly destroyed but one wing is complete. The second paratype has only one wing and fragments of the body preserved.
Etymology. Named after M. Jean-Bernard Myskowiak, father of the first author.
Type locality. Farm Le Quesnoy, 49°19'31" N, 2°40'22" E, Chevrière, Region of Creil, Oise Department (northern France).
Type horizon. Lowermost Eocene amber, Sparnacian (53 Ma), level MP7 of the mammal fauna of Dormaal ( Brasero et al. 2009).
Diagnosis As for the genus.
Descriptions.
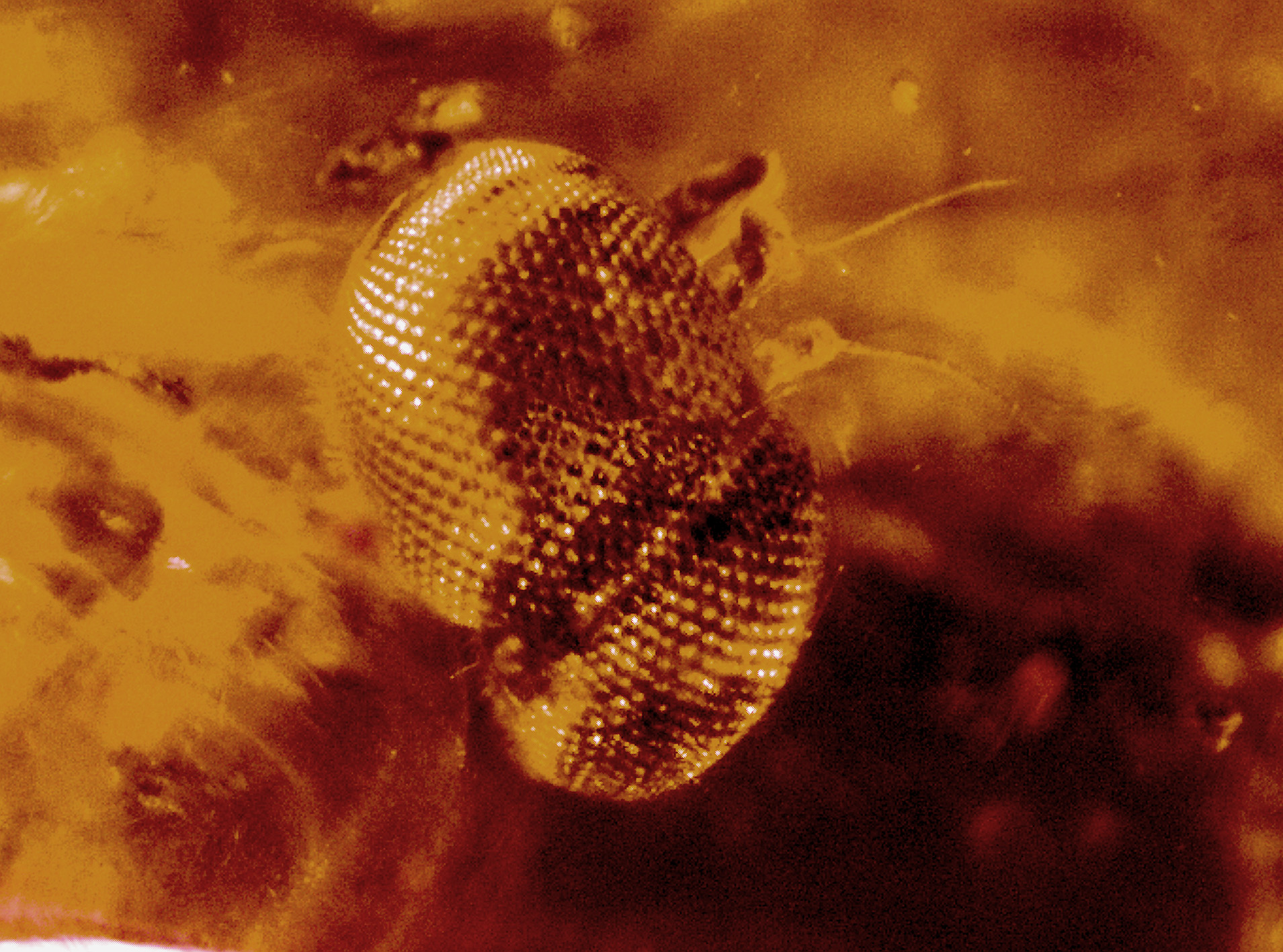
Head: female ( Figs 1–2 View FIGURE 1 View FIGURE 2 ) with only its dorsal surface is preserved with antennae, head 30.84 mm long, 1.18 mm wide; eyes well separated, 0.34 mm apart, less than half of their width; front just above antennae distinctly narrower than one eye, provided with rather long hairs; three ocelli in triangle on vertex; antennae hardly visible, with bases contiguous, with distance between their base clearly less than diameter of median ocellus, scape and pedicel globose, basal flagellomere extending ventrally beyond pedicel; arista subapical on basal flagellomere, narrow and elongate; male head holoptic, eyes meeting dorsally; male gena provided with numerous long setae; face not visible.
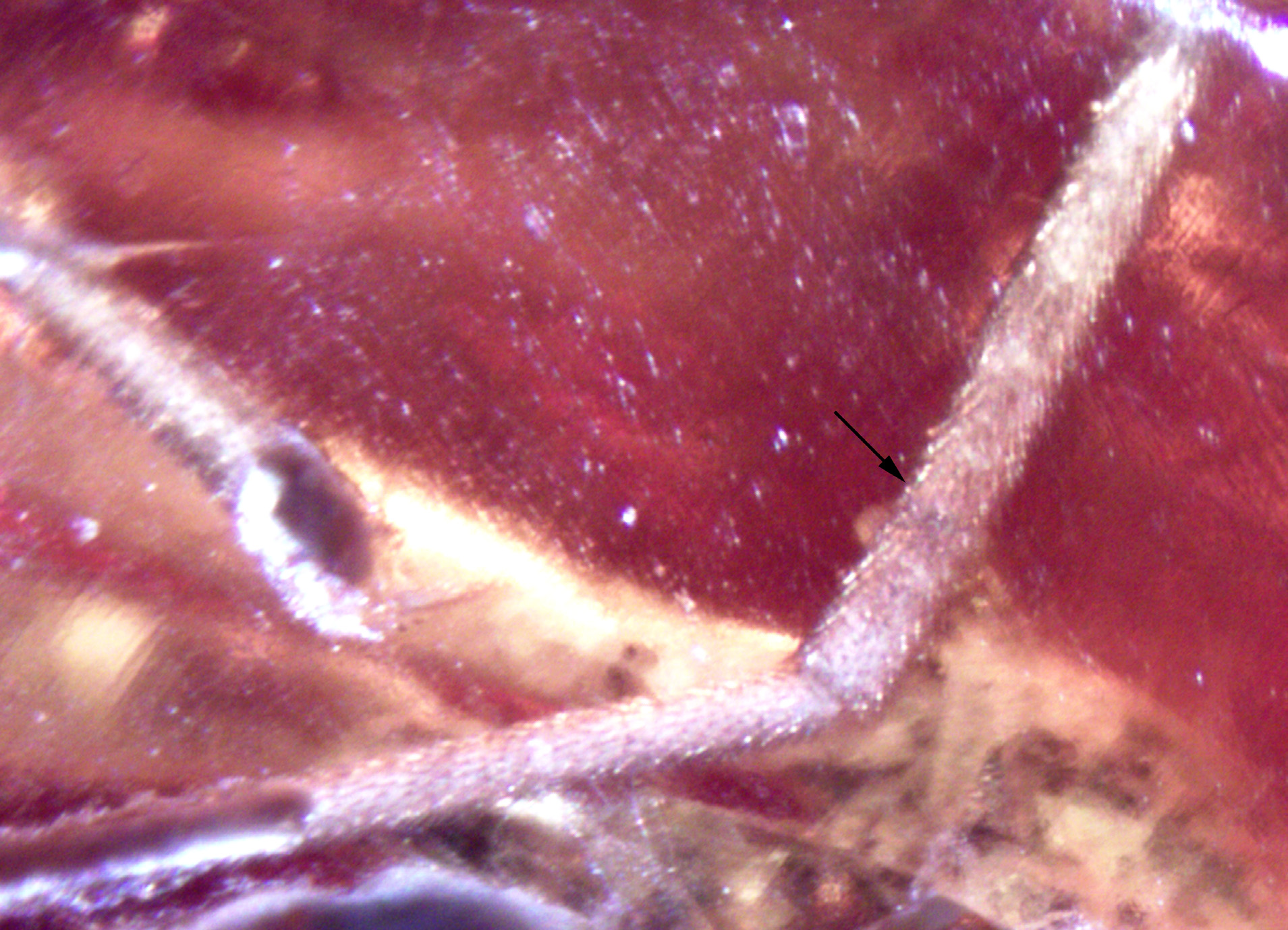
Thorax: ( Fig. 3 View FIGURE 3 ) 1.9 mm long, 1.3 mm wide, with white pair of white stripes divided medially by broad dark stripe and two darker spots on sides of thorax; with numerous short erect fuscous setae arranged in no distinct pattern; sclerite between anterior spiracle and fore coxa with knob-like ( Fig. 4 View FIGURE 4 ) process; subscutellum not visible, hidden below large and broad scutellum, 0.35 mm long and 0.76 mm wide; posterior thoracic spiracle with a scalelike elevation immediately behind.
Wing: ( Fig. 5 View FIGURE 5 ) hyaline, relatively large and broad, 4.39 mm long, 1.5 wide, broadest basally near basal end of large anal lobe; apex of wing moderately rounded; alula and lower calypter large; vein R2+3 ending very near to R1; vein R4 terminated at wing apex; veins M1, M2, and M3 shorter than cell dc; basal radial cell reaching level of apex of Sc; vein R4 gently arched; cell dc 1.07 mm long, 0.26 mm wide; cell m3 open at margin but with M3 and CuA1 strongly convergent; anal cell closed but CuA2 and A1 touching only at wing margin.
Legs: ( Fig. 6 View FIGURE 6 ) Fore and middle legs slender; fore tarsus with long setae below; femur laterally flattened except basally; tibia strongly flattened; tibial spur formula 0/2/2 (see below for paratype PA 9055 2/6) ( Figs 7–8 View FIGURE 7 View FIGURE 8 ). Fore femur 1.36 mm long, mid femur 1.25 mm long, tibia 1.41 mm long, tarsus 1.36 mm long; hind femur 1.63 mm long, tibia 1.74 mm long, tarsus 0.81 mm long.
Abdomen: female abdomen proportionately short, 3.17 mm long, 1.18 wide; slightly broader than thorax, black with large sclerotized tergites covered with small to medium setae; T1 largest segment; T2–5 about equal in lengths, T6 and T7 shorter, T8 shortest (only about half length of T7); cerci one-segmented ( Fig. 9 View FIGURE 9 ), covered with very short dense setae; male abdomen narrower than in female with genitalia partly visible but difficult to interpret. Paratype female PA 9055 2/6 ( Figs 8 View FIGURE 8 , 11–12 View FIGURE 11 View FIGURE 12 ). Its attribution to the same species is based on the identity of pattern of the body and wing venation as for the female allotype. Same as allotype, with fore leg well preserved, lacking tibial spurs; mid and hind tibiae with pair of spurs.
Discussion. These fossil flies belong to the same species because of the wing venation and body patterns of coloration of the different male and female specimens. The male is holoptic, unlike the females, which is a dimorphic trait frequent among flies, and especially in Athericidae . Their presence in the same small piece of amber also supports their attribution to the same species. The wing venation of Eoatrichops gen. n. is of the type present in Athericidae but also in some Xylophagidae . The subscutellum is not visible, hidden below the scutellum, which renders it inaccessible, and is characteristic of the Athericidae . Nevertheless the flagellum with a slender nonannulated subapical arista is a character present in Athericidae , while Xylophagidae have flagellum with several flagellomeres or an arista in apical position. The female one-segmented cerci is a putative synapomorphy of the ( Athericidae + Tabanidae ) (after Kerr 2010). A further character is the presence of a scale-like elevation immediately behind posterior thoracic spiracle, present in Athericidae , Tabanidae , and Pelecorhynchidae but not in Xylophagidae (after McAlpine 1981). The apical parts of veins R2+3 and R1 very close is diagnostic and synapomorphic for the Athericidae . In fact, according to Kerr (2010) the synapomorphy would be R2+3 and R1 touching at wing margin, but some modern Athericidae (e.g. Suraginella Stuckenberg, 2000 ) have these veins extremely close as in our fossil but not exactly touching ( Stuckenberg 2000).
Following the key to genera of modern Athericidae of Nagatomi (1985), Eoatrichops would fall near the genus Atrichops for the following characters: Antennal segment 3 reniform with dorsal arista; hypopleura with a pit just above hind coxa; antennae almost contiguous with each other; sclerite between anterior spiracle and fore coxa with one knob-like process at lower margin [propleural projection above fore coxa sensu Stuckenberg (1974)]. Nevertheless, Eoatrichops differs from Atrichops in the tibial spur formula (0/2/ 2 in Eoatrichops while it is 0/1/ 2 in Atrichops ) ( Nagatomi 1979, 1984). A further difference would be the absence of a short fusion of A1 with CuA 2 in Eoatrichops while it is present in the modern Atrichops . Stuckenberg (1974) considered that both the tibial formula and the presence of a long-stalked anal cell are characters important to recognize the genus Atrichops . Nevertheless the last character seems to be rather variable within a given genus of Athericidae because a stalked anal cell is present in some Atherix Meigen, 1803 and absent in others (see Stuckenberg 1973; Webb 1981). Eoatrichops differs from the Latest Eocene-Earliest Oligocene Atrichops hesperius Cockerell, 1914 , unique fossil currently attributed to this genus, in the absence of a short fusion A1+CuA2 ( Cockerell 1914).
Among the modern athericid genera described after 1985, Suraginella Stuckenberg, 2000 shares with Eoatrichops the antennal bases less distant than the diameter of median ocellus, and shape of pedicel, but it differs from it in the cell m3 closed ( Stuckenberg 2000). The genus Asuragina Yang & Nagatomi, 1992 strongly differs from Eoatrichops in the widely separated bases of the antennae ( Yang & Nagatomi 1992). Among the fossil athericid genera, Eoatrichops differs from the Baltic amber Succinatherix Stuckenberg, 1974 in the tibial formula 0/2/2 instead of 1/2/2 and the absence of the propleural projection, even if they share the close antennal bases and reniform antennal segment 3 ( Stuckenberg 1974). Eoatrichops differs from the Lower Cretaceous genus Athericites Mostovski et al., 2003 in the basal radial cell reaching the level of apex of Sc instead of being much shorter, and vein R4 not sigmoidal ( Mostovski et al. 2003). Eoatrichops differs from the Lower Cretaceous genus Sinocretomyia Zhang, 2012 in the vein R2+3 nearly straight instead of being strongly sigmoidal and the arista subapical instead of being apical on basal flagellomere ( Zhang 2012). The Lower Cretaceous Palaepangonius Ren, 1998 differs from Eoatrichops in the different shaped discal cell, with proximal part distinctly narrower than distal part, and vein R4 much more sigmoidal ( Ren 1998). The mid Cretaceous Galloatherix is characterized by the veins M1, M2 and M3 as long as discal cell, unlike Eoatrichops ( Nel et al. 2014).
Note. Atherix saunieri Théobald, 1937 (Late Eocene of Gard department, France, type stored in the Musée of Nimes, could not located, and probably lost) has a vein R2+3 ending on anterior wing margin very far from apex of R1, which is clearly not a character for Athericidae (see Théobald 1937: pl. 11, fig. 7). Its venation more strongly resembles that of a Rhagionidae . We consider that it does not belong to the Athericidae and it is a Diptera of uncertain family, nov. stat.











| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |