
Envekadea vanlandinghamii Graeff, Kociolek & S.R. Rushforth, 2013
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.153.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/546B8784-F961-397A-FF3D-7F19FA8E60C9 |
|
treatment provided by |
Felipe |
|
scientific name |
Envekadea vanlandinghamii Graeff, Kociolek & S.R. Rushforth |
| status |
sp. nov. |
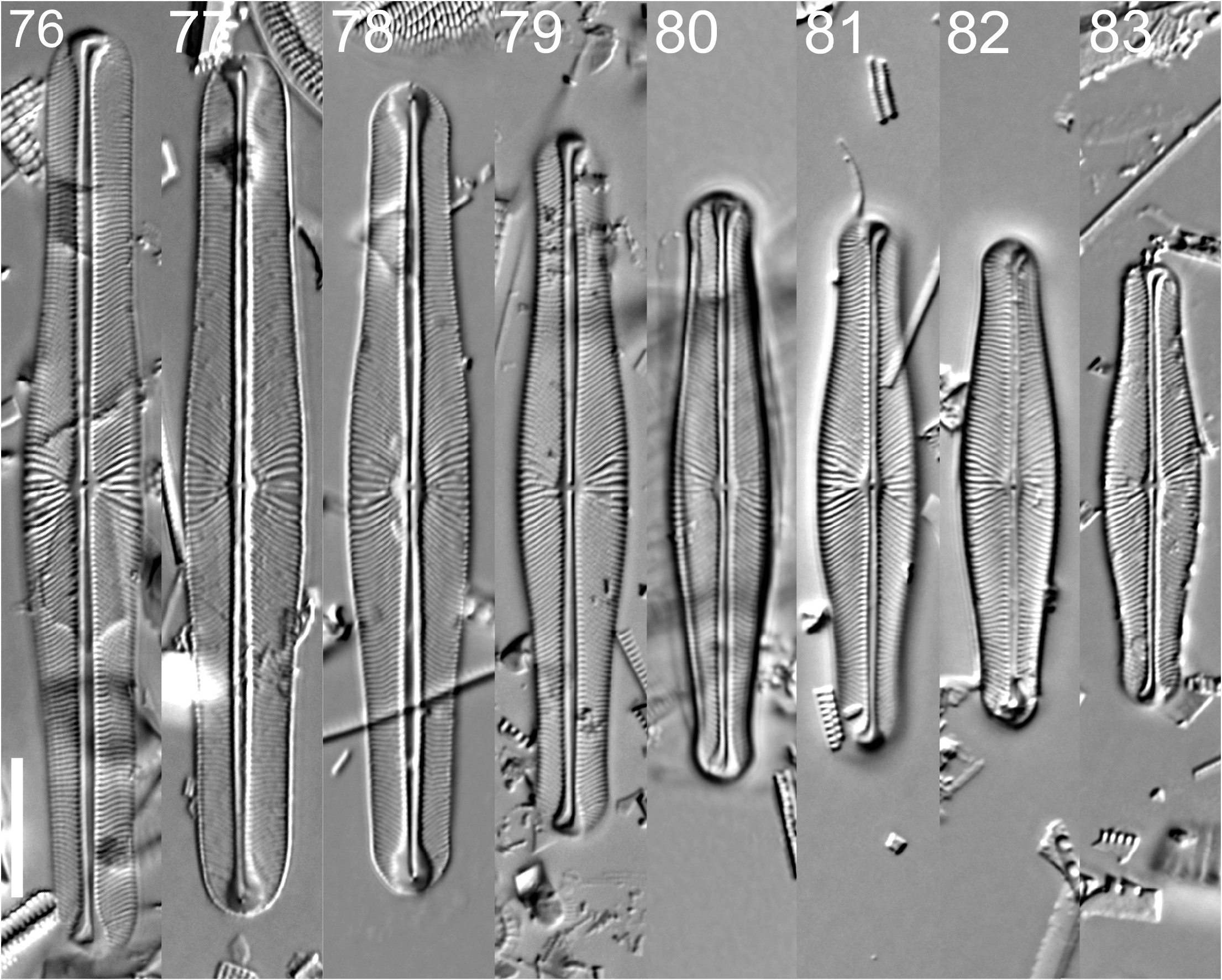
Envekadea vanlandinghamii Graeff, Kociolek & S.R. Rushforth , sp. nov. ( Figs 76–89 View FIGURES 76–83 View FIGURES 84–89 ; figure 79 =
holotype)
Valves linear, tumid in the center, with apices protracted, rounded ( Figs 76–83 View FIGURES 76–83 ). Length 32.0–67.0 µm, breadth 7.0– 8.5 µm. Apical breadth 4.0–6.0 µm. Axial area wide at the ends of the valve, very narrow to the center where it is only slightly wider and bordered on either side by striae that are more coarse than the others and of various lengths. The raphe is weakly lateral with external proximal ends terminating close to one another and distal ends deflected in opposite directions ( Figs 76, 79, 81 View FIGURES 76–83 ). Striae are indistinctly punctate, strongly radiate around the center, strongly convergent towards the poles ( Figs 79, 81 View FIGURES 76–83 ). Striae 17–23/ 10 µm in the center, 27–29/ 10 µm at the ends.
In the SEM, the valve exterior is dominated by the striae with their volate occlusions ( Fig. 85 View FIGURES 84–89 ) and a sharp change in orientation from radiate at the center to convergent at the ends ( Fig. 84 View FIGURES 84–89 ). The raphe branches and axial area are S-shaped ( Fig. 84 View FIGURES 84–89 ). Proximal raphe ends are widely T-shaped ( Fig. 86 View FIGURES 84–89 ). The terminus of the axial area is wide and more developed on one side than the other ( Fig. 85 View FIGURES 84–89 ). Internally, the axial area is narrow and thickened ( Fig. 87 View FIGURES 84–89 ). Helictoglossae are fine, with the end pointed inward ( Fig. 88 View FIGURES 84–89 ). Proximal raphe ends terminate close to one another ( Fig. 89 View FIGURES 84–89 ).
Type:— USA. Blue Lake , Tooele Co., Utah. ( COLO 439035 About COLO , holotype! (= Fig. 79 View FIGURES 76–83 ), designated here; COLO 8506 About COLO , BM 101680, isotypes) .
Etymology:—Named in honor of Dr. Sam VanLandingham, who passed away in 2013, for his many contributions to the field of diatom research.
Distribution:— Envekadea vanlandinghamii can be found in the main basin of Blue Lake in varying abundance.
Observations:—This is the first report of this genus in USA inland waters. Two other known species in the genus, E. pseudocrassirostris (Hustedt) Van de Vijver et al. in Gligora et al. (2009: 136) and E. hedinii (Hustedt) Van de Vijver et al. in Gligora et al. (2009: 137), originally described from Norway and Tibet, respectively, are morphologically very similar to E. vanlandinghamii ( Gligora et al. 2009) , but can be distinguished by their differing valve outlines and central area size. E. vanlandinghamii has a slightly larger central area than E. pseudocrassirostris and valve margins that widen at the apices rather than taper. E. vanlandinghamii has valves that are less strongly undulate than those of E. hedinii ( Simonsen 1987: plate 87, figs 5–8). A recently described species of the genus, E. metzeltinii Lee et al. , can be distinguished from other known species in the genus by the presence of longitudinal ribs and relatively parallel valve margins ( Lee et al. 2013).
Envekadea is geographically widespread in the Northern Hemisphere, with reports from eastern Asia, across Europe, eastern North America ( Lee et al. 2013; Gligora et al. 2009), and now central North America. E. pseudocrassirostris , or Navicula pseudocrassirostris Hustedt , has been reported in North America from Pettaquamscutt River, Rhode Island ( Kennett and Hargraves 1985) and Biscayne Bay, Florida ( Wachnicka et al. 2011).
An image of a specimen of Envekadea identified by Witkowski et al. (2000: pl. 157, fig. 4) as N. pseudocrassirostris Hustedt (1961: 79) , presumably from a locality in Europe, looks much like E. vanlandinghamii with its slightly larger central area and noticeably broader apices. This suggests that E. vanlandinghamii may not be restricted to central North America.
Not only is Envekadea geographically widespread in the Northern Hemisphere, it is also ecologically pervasive. E. vanlandinghamii and E. pseudocrassirostris have been collected from warm mesohaline habitats in North America ( Wachnicka et al. 2011: table 1; personal observations). Navicula hedinii Hustedt (1922a: 132) was originally described from a freshwater lake ( Gligora et al. 2009: 124) and additional specimens come from a high conductivity, low salinity lake in Croatia ( Gligora et al. 2007). Witkowski et al. (2000: pl. 156, fig. 17) reported a specimen of Envekadea , which has a valve outline similar to that of E. hedinii , from the high salinity Adriatic Sea ( Artegiani et al. 1997). Witkowski et al. (2000: pl. 157, fig. 5) also collected N. pseudocrassirostris from Lago di Varano in Italy, a coastal pond that has a surface salinity range of 17 to 27 ppt ( Spagnoli et al. 2002). E. metzeltinii was described from coastal habitats in Florida and also reported from the Yucatan region of Mexico ( Lee et al. 2013).
Fallacia pseudolitoricola (Håkansson) Håkansson in Snoeijs & Vilbaste (1994: 45) ( Figs 90–105 View FIGURES 90–96 View FIGURES 97–100 View FIGURES 101–105 ) Basionym: Navicula pseudolitoricola Håkansson (1982: 90 , pl. 1, figs 4–7, pl. 2, figs 8–12)
Valves are linear-elliptical with rounded apices ( Figs 90–96 View FIGURES 90–96 ). Length 26.0– 42.5 µm, breadth 9–11 µm. The axial area is narrow, slightly expanded then constricted near the center. No discernible central area is present. The raphe is weakly lateral, straight to slightly arched, with the external distal raphe ends deflected in the same direction onto the mantle ( Figs 90, 96 View FIGURES 90–96 ). A relatively wide canal running longitudinally is located on either side of the axial area, which is bordered by a narrow row of areolae ( Figs 91, 96 View FIGURES 90–96 ). The striae are radiate, more strongly so near the apices, and distinctly punctate ( Fig. 93 View FIGURES 90–96 ). Striae number 20–21/ 10 µm.
In the SEM, the valve exterior has a wide conopeum extending over the valve face for about ¼–⅓ of the area ( Fig. 97 View FIGURES 97–100 ). The conopeum has on either side of the raphe one to two rows of irregularly-arranged small depressions ( Figs 98– 100 View FIGURES 97–100 ). These probably open internally into the canal. At the outer edges of the conopeum there is the edge of the longitudinal canal or depression seen with areolae along that edge ( Figs 98, 100 View FIGURES 97–100 ). The striae are comprised of rounded areolae ( Figs 97–100 View FIGURES 97–100 ). At the ends, the raphe curves onto the mantle ( Figs 97–99 View FIGURES 97–100 ). Internally, the interior of the valve has longitudinal canals superficially lyre-shaped ( Fig. 101 View FIGURES 101–105 ). The canals can extend to the center of the valve ( Fig. 105 View FIGURES 101–105 ) or stop short of the center ( Figs 101, 103 View FIGURES 101–105 ). The proximal raphe ends terminate close to one another ( Fig. 103 View FIGURES 101–105 ), and at the distal ends the helictoglossae are small ( Fig. 102 View FIGURES 101–105 ). No coverings of the areolae have been discerned internally or externally ( Figs 97–105 View FIGURES 97–100 View FIGURES 101–105 ).
Distribution:— Fallacia pseudolitoricola can be found rarely in the main basin of Blue Lake and more abundantly in Chara periphyton of the marsh on the west side of Blue Lake (COLO 8523).
Observations:—Prior to the description of Fallacia A.J. Stickle & D.G. Mann in Round et al. (1990: 667) specimens of this taxon were considered part of the genus Diploneis (Ehrenberg) Cleve (1894: 76) . Fallacia pseudolitoricola has been confused with Diploneis in the light microscope due to similar valve outlines, stria appearances, and pseudo-lyre-shaped structures. Kaczmarska & Rushforth (1983) identified specimens of F. pseudolitoricola as Diploneis marginestriata Hustedt (1922b: 236) . A later report of D. marginestriata made by Squires & Rushforth (1986) from the “mildly saline” Utah Lake, located roughly 150 km northwest from Blue Lake, suggests that F. pseudolitoricola has also been found in that body of water. Idei & Kobayasi (1988: 278, fig. 1) and Krammer & Lange-Bertalot (1986: fig. 110: 4) contribute light micrographs from the type population of D. marginestriata , which show specimens with striae that are not obviously punctate as they are in F. pseudolitoricola and internal costae that continue below the hyaline region along the external axial area. Comparisons of the taxa using scanning electron microscopy reveal very different valve structures and confirm that F. pseudolitoricola is not a member of Diploneis ; the presence of a conopeum and the absence of costae distinguish Fallacia from Diploneis ( Round et al. 1990) . In addition, F. pseudolitoricola does not belong under the marine genus Lyrella Karayeva (1978: 1595) , which is also known for having a large lyre-shaped structure. In Lyrella a conopeum is lacking, and the lyre-shaped structure is a solid thickening of silica instead of a canal ( Round et al. 1990).
Håkansson (1982: 90) originally described Navicula pseudolitoricola from the Baltic Sea. In a later publication, in which she transfers the taxon to Fallacia, Håkansson (in Snoeijs & Vilbaste 1994: 45) notes that the species has only been found in fossil sediments. Our report documents a live collection of the species. F. pseudolitoricola has also been collected live in the Bahamas from a mangrove pond but we believe it has been identified incorrectly as F. litoricola (Hustedt) Mann in Round et al. (1990: 668; basionym: Navicula litoricola Hustedt 1955: 23 , pl. 8, figs 13–14) in Hein et al. (2008: pl. 25, fig. 5). Our specimens of F. pseudolitoricola are slightly smaller and less finely striate than the type population which is 28.0–42.0 µm long and 9.5–13.5 µm wide and has a stria density of 22–26/ 10 µm ( Håkansson 1982).
There are a few taxa that are morphologically similar to F. pseudolitoricola . F. schaeferae (Hustedt) Mann in Round et al. (1990: 669, basionym: Navicula schaeferi (schaferi) Hustedt 1964: 545, fig. 1583) was originally described from Wangeroog, Germany ( Simonsen 1987: 491, pl. 751, figs 20–22, Hustedt 1964: 545, fig. 1583) and also reported from the Adriatic Sea ( Witkowski 2000: 212, pl. 70, fig. 29). Håkansson (1982) explored the similarities between F. pseudolitoricola and F. schaeferae using light microscopy and notes that the striae of F. schaeferae are finer and that an uninterrupted row of areolae is adjacent to either side of the raphe. Images provided by Pankow (1976) and Cleve-Euler (1953) suggest that the Northern European taxa Navicula plathi Brockmann (1950: 19) and N. hyalina Donkin (1861: 10) are also morphologically similar to F. pseudolitoricola , and differences in the widths of the conopea relative to the valves distinguish these taxa from F. pseudolitoricola . More recently, N. plathi ( plathii ) Brockmann (1950: 19, pl. 3, figs 25–27) has been transferred to Fallacia by Snoeijs as Fallacia plathii (Brockmann) Snoeijs (in Snoeijs & Balashova 1998: 50, fig. 438) and N. hyalina Donkin (1861: 10 , pl. 1, fig. 6 non Navicula hyalina Hajós 1968: 173 , pl. 52, fig. 10) was transferred to Diploneis by Cleve (1894: 80). F. pseudolitoricola is morphologically distinct from several other brackish or marine Fallacia from North American and Northern European regions based on valve size, appearance of the striae, and shape of the canal and/or conopeum ( Sabbe et al. 1999, Witkowski 1991).




| BM |
Bristol Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Envekadea vanlandinghamii Graeff, Kociolek & S.R. Rushforth
| GRAEFF, C. L., KOCIOLEK, J. P. & RUSHFORTH, S. R. 2013 |
Fallacia pseudolitoricola (Håkansson) Håkansson
| Snoeijs, P. & Vilbaste, S. 1994: 45 |
| Hakansson, H. 1982: 90 |