
Enhydrosoma robustum, Karanovic, Tomislav, Kim, Kichoon & Lee, Wonchoel, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.3990.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:90BE4EAF-8594-4164-AE26-4F47E04A4A5D |
|
DOI |
https://doi.org/10.5281/zenodo.5616524 |
|
persistent identifier |
https://treatment.plazi.org/id/03CE8781-FFC2-E64B-6180-2B63FB3EE4B7 |
|
treatment provided by |
Plazi |
|
scientific name |
Enhydrosoma robustum |
| status |
sp. nov. |
Enhydrosoma robustum sp. nov.
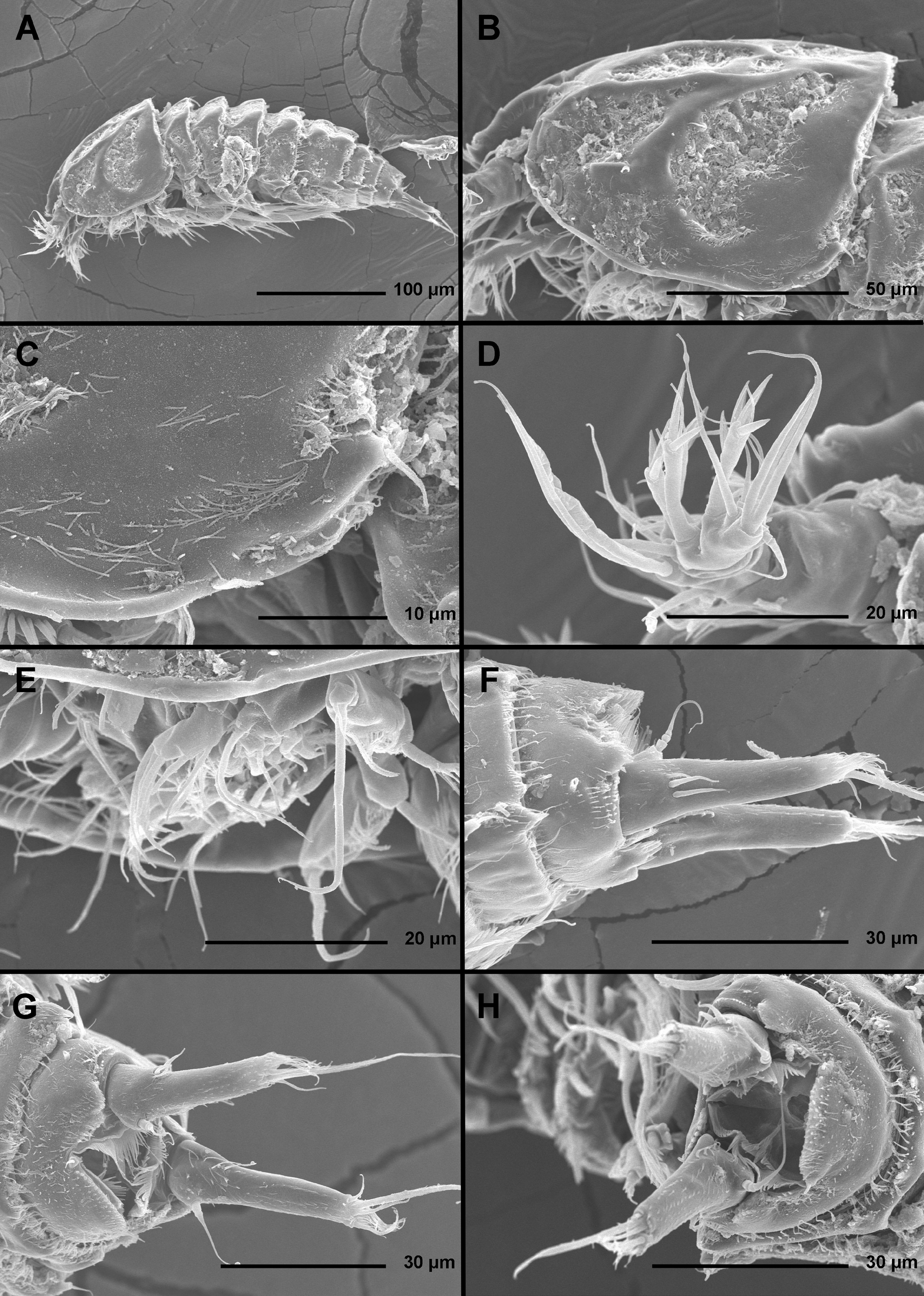
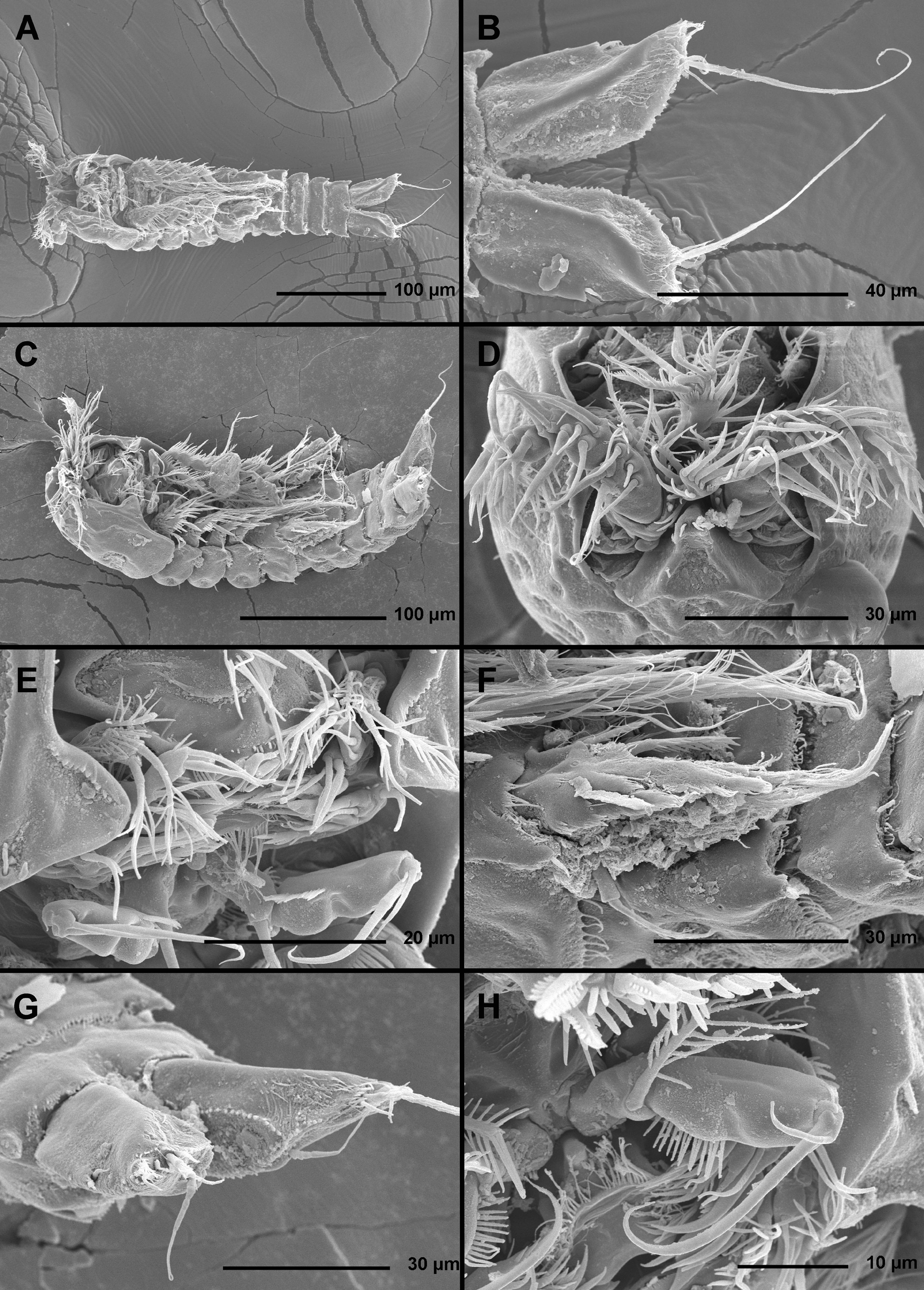
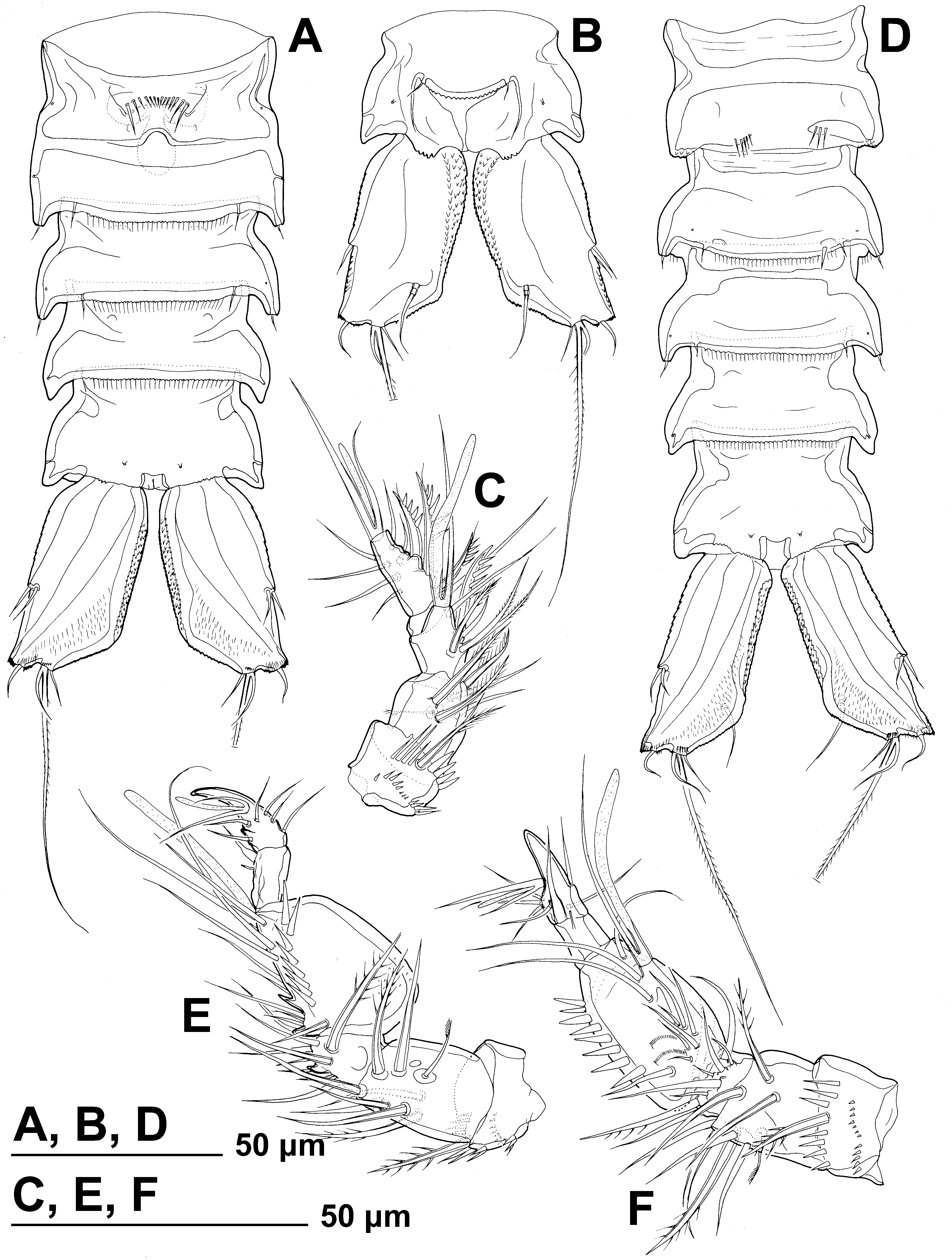
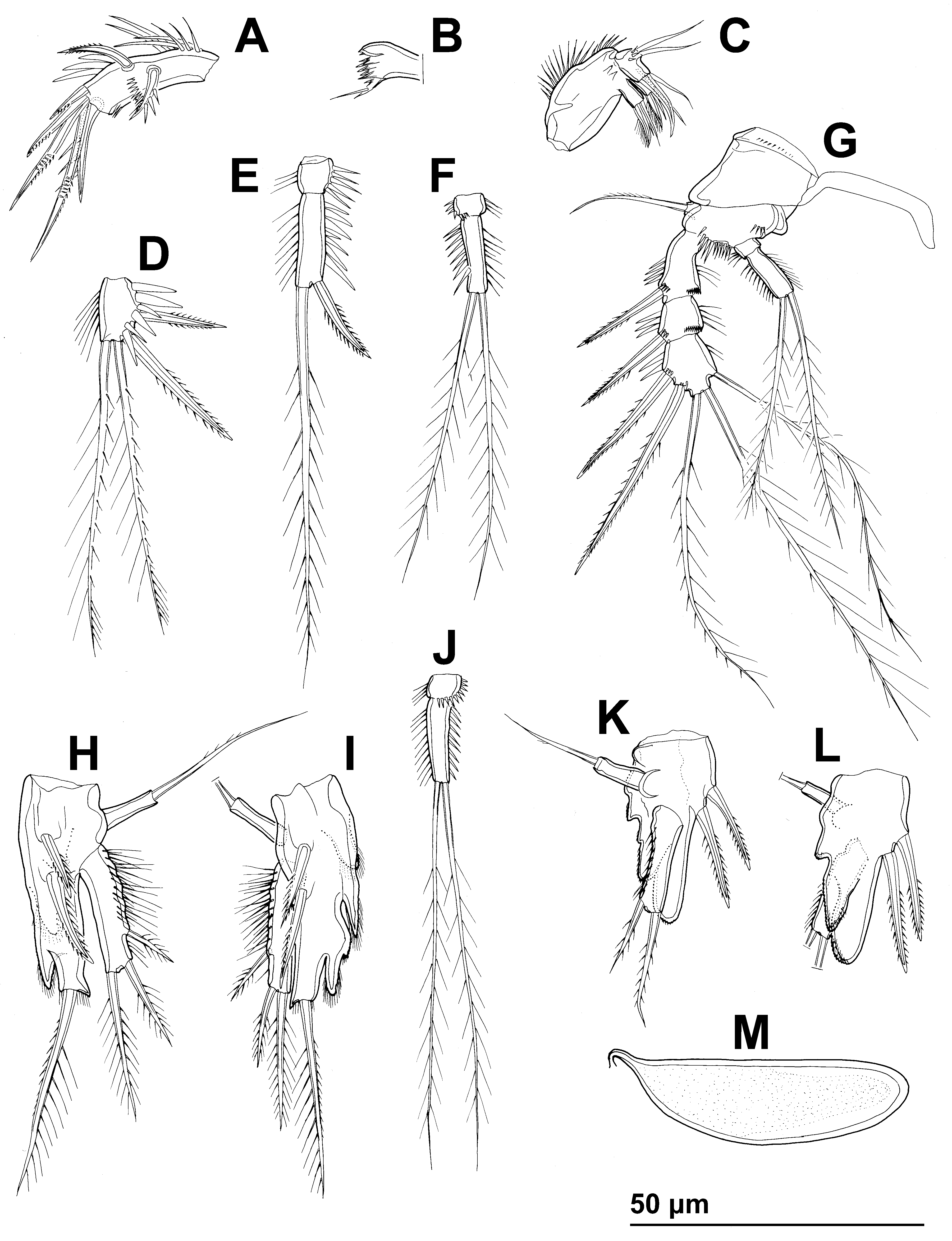
( Figs. 10–15 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 )
Type locality. South Korea, West Coast, Yellow Sea, Garorim Bay, muddy sediments, N 3658'32.3" N 12619 View Materials '17.9"E.
Specimens examined. Holotype female (NIBRIV 0000287216) preserved in 70% ethanol collected from the type locality, 1 November 2012, collected by K. Kim. Dissected allotype male (NIBRIV 0000287217) mounted on one slide, collected from Korea, South Sea, Gwangyang Bay, sampling station 12 (see Karanovic et al. 2014), 34°53'24.50"N 127°47'42.40"E, 25 January 2006, collected by K. Kim. Additional paratypes: one female (NIBRIV 0000287218) preserved in 70% ethanol collected from the type locality; three dissected females (NIBRIV 0 0 0 0 287219, NIBRIV 0 0 0 0 287220, NIBRIV 0000287221) mounted on one slide each, collected from the type locality by K. Kim, 1 November 2012; one male and four females together on one SEM stub (NIBRIV 0000287222), collected from the type locality by K. Kim, 1 November 2012; three males and three females together on another SEM stub (NIBRIV 0000287223) collected from Korea, South Sea, Gwangyang Bay, sampling station 12 (see Karanovic et al. 2014), 34°53'24.50"N 127°47'42.40"E, 25 January 2006, collected by K. Kim. One female used for molecular analysis collected by K. Kim from the type locality on 1 November 2012 (see Table 1 View TABLE 1 ).
Etymology. The species name derives from the Latin adjective robustus, meaning “hard and strong like oak”, and refers to the robust shape of the caudal rami. It was treated as an adjective, agreeing in gender with the neuter generic name.
Description of female. Based on holotype and several paratypes. Total body length from 287 to 342 µm (mean = 318 µm, n = 12). Body segmentation, colour, nauplius eye, hyaline fringes, general integument thickness, and most somite ornamentation as in Enhydrosoma apimelon sp. nov. However, surface relief on most somites different, and angle between telescopic and non-telescopic part of pleurons on most free somites much sharper (compare Figs. 1 View FIGURE 1 A and 10A). Habitus ( Figs. 10 View FIGURE 10 A, G, 11A, C, 12A, C, E) generally cylindrical in dorsal view, widest at posterior end of cephalothorax and tapering posteriorly, boundary between prosome and urosome inconspicuous; prosome/urosome length ratio about 1.4, and prosome only slightly more voluminous than urosome. Body length/ width ratio about 3.8 in dorsal view; cephalothorax about 1.4 times as wide as genital double-somite. Free pedigerous somites without lateral or dorsal expansions, heavily sculptured, pleurons only partly covering coxae of swimming legs in lateral view. Integumental relief more defined by depressions than ridges in dorsal and lateral view; ventro-lateral ridges on urosomites as in E. apimelon . Most cuticular depressions and posterior margin of somites without hair-like spinules (except ventrally on urosomites), but full of bacterial growth and detritus, hindering observation of cuticular pores and sensilla. Hyaline fringe of all somites narrow and rough, in some places nearly serrated. In addition to hair-like spinules, surface ornamentation of somites and caudal rami consists of at least two different types of sensilla (slender and bottle-shaped; see Fig. 10 View FIGURE 10 D, F), simple cuticular pores, tubular pores, and few stronger spinules; exact number of pores and spinules difficult to establish.
Rostrum ( Figs. 10 View FIGURE 10 B, 11D, 13A) as in E. apimelon , except for space between sensilla smaller than one horn-like projection.
Cephalothorax ( Figs. 10 View FIGURE 10 B, C, D, 11D, E) shape as in E. apimelon , but with completely different relief, without comb of long setules in anterior part along interior surface of lateral margin ( Fig. 11 View FIGURE 11 D), instead ventral margin produced ventrally into two characteristic flaps that protect first three pairs of mouth appendages ( Fig. 11 View FIGURE 11 E), and distal part of lateral margin finely serrated ( Fig. 10 View FIGURE 10 C). Dorsal surface depressions more elongated than in E. apimelon (compare Figs. 3 View FIGURE 3 B and 10G), and lateral depressions smaller (compare Figs. 1 View FIGURE 1 B and 10B), but most sensilla easy to homologize.
Pleuron of free prosomites ( Figs. 10 View FIGURE 10 E, 12A) very similar to each other, lacking triangular dorsal plates, conical mound-looking protuberances at base of sensilla, and posterior hair-like spinules; with oval dorsal and semicircular lateral depressions. H higher areas of relief not defined into ridges and surface less smooth; in addition to sensilla homologous to those in E. apimelon , each free prosomite with one anterior dorso-lateral pair of small pores, and one central dorsal pair of large pores.
First urosomite ( Figs. 10 View FIGURE 10 A, G, 12A) slightly longer and narrower than fourth pedigerous somite and pleuron without free lateral margin, but relief very similar to that of other pedigerous somites.
Genital double-somite ( Figs. 10 View FIGURE 10 G, 11A, F, 12A, C, E, 14A) 1.3 times as wide as long in ventral view; completely fused ventrally but with deep suture indicating original segmentation between genital and third urosomites dorsally, thus dividing double-somite into equally long and similarly wide halves; ventral surface relatively smooth and flat, with ornamentation as in E. apimelon and with ventro-lateral ridges pronounced and produced posteriorly with pore on tip ( Fig. 14 View FIGURE 14 A). Dorsal and lateral surfaces with relief, but without pronounced ridges and posterior protrusions; female genital complex ( Fig. 14 View FIGURE 14 A) weakly sclerotized and hardly distinguishable from internal sutures and soft tissue, except for large copulatory pore near midlength of somite and wide copulatory duct; genital operculum as in E. apimelon .
Third urosomite ( Figs. 10 View FIGURE 10 G, 11A, F, 12A, C, E, 14A) very similar in shape and ornamentation to posterior part of genital double-somite, with flat ventral surface, large posterior projections of ventro-lateral ridges with sensilla on tip, pair of ventral posterior sensilla, posterior row of ventral and lateral hair-like spinules, ventro-lateral pores, and two posterior dorsal pairs of sensilla; no dorso-lateral ridges, with nearly semi-circular shape in cross-section.
Fourth urosomite ( Figs. 10 View FIGURE 10 G, 11A, C, G, 12A, C, E, G, 14A) shorter and narrower than third urosomite, with shorter posterior projections of dorso-lateral and ventro-lateral ridges, without sensilla, ornamented with pair of ventro-lateral pores, and posterior row of long hair-like spinules on ventral and partly on lateral surface.
Anal somite ( Figs. 10 View FIGURE 10 H, 11G, 12G, 14A, B) only slightly clefted medially, with one pair of large dorsal sensilla at base of anal operculum, one pair of dorso-lateral simple pores, one pair of lateral simple pores, and one pair of ventral simple pores; ventro-lateral corners produced as in preanal somite, but slightly shorter. Anal operculum ( Figs. 10 View FIGURE 10 H, 14B) semi-circular, short, only reaching slightly beyond midlength of somite, serrated, representing 43% of somite's width; anal sinus ( Figs. 10 View FIGURE 10 H, 14B) widely open, without hair-like spinules.
Caudal rami ( Figs. 10 View FIGURE 10 H, 11B, G, 12B, G, 14A, B) long, stout, about twice as long as anal somite, widest in central part, with narrower base and only tapering posteriorly at distal quarter, about twice as long as wide (ventral view), slightly divergent, with strong dorsal ridge giving near triangular cross-section, with very small space between them. Anterior part of medial margin flat; armature as in E. apimelon but dorsal and anterior lateral setae inserted much more posteriorly (at about 3/5 of ramus’ length). Ornamentation consisting of several hair-like spinules on ventral side posteriorly and at base of distal lateral seta, and small spinules along ridges; no tubular pore at base of anterior lateral setae; outer distal process carrying posterior lateral seta even more produced than in E. apimelon ; proportion of setae similar to that in E. apimelon ; principal apical seta 1.3 times as long as caudal rami.
Antennula ( Figs. 11 View FIGURE 11 D, 12D, F, H, 13A, 14C) short, stout, and five-segmented, as in E. apimelon . Lengthof segments and most armature as in E. apimelon , except for second segment with only seven setae, thus armature formula: 1.7.7+ae.1.11+ae; as in E. apimelon one pinnate seta on third segment and all three pinnate setae on fifth segment recurved, spiniform, with very strong spinules only along convex surface.
Antenna ( Figs. 11 View FIGURE 11 D, 12D, F, 15A), labrum ( Fig. 11 View FIGURE 11 E), paragnaths ( Fig. 11 View FIGURE 11 E), mandibula ( Figs. 11 View FIGURE 11 E, 15B), maxillula ( Fig. 11 View FIGURE 11 E), maxilla ( Figs. 11 View FIGURE 11 E, 15C), maxilliped ( Fig. 11 View FIGURE 11 E, H), first swimming leg ( Fig. 15 View FIGURE 15 D, E), second swimming leg, third swimming leg ( Fig. 15 View FIGURE 15 F), and fourth swimming leg ( Fig. 15 View FIGURE 15 G) as in E. apimelon , except for small differences in proportion of certain segments and armature elements and very minor differences in ornamentation.
Maxilla ( Fig. 15 View FIGURE 15 C) without tubular pore and inner row of spinules.
Endopod of third swimming leg ( Fig. 15 View FIGURE 15 F) and especially endopod of fourth swimming leg ( Fig. 15 View FIGURE 15 G) proportionately shorter than in E. apimelon , and all exopodal segments on all swimming legs also slightly shorter.
Fifth leg ( Figs. 11 View FIGURE 11 F, 15H, I) biramous as in E. apimelon , comprising conical exopod and baseoendopod, but exopod slightly shorter than endopodal lobe; exopod also armed with three elements, but lateral elements inserted closer to distal margin than in E. apimelon , with distal lateral element almost subapical and difficult to homologize; outer margin of exopod with numerous long hair-like spinules, inner margin smooth. Baseoendopod armed with two inner strong spines and one distal seta as in E. apimelon , but distal seta much longer and very strong; inner margin of baseoendopod with three spiniform projections, each with tuft of hair-like spinules, and no tubular pores. Distal exopodal element about 2.2 times as long as proximal endopodal spine, 1.5 times as long as distal endopodal spine, 2.9 times as long as proximal exopodal element, 1.6 times as long as middle exopodal element, 0.7 times as long as distal endopodal element, and nearly 1.2 times as long as entire exopod.
Sixth legs ( Fig. 14 View FIGURE 14 A) as in E. apimelon , but with relatively smaller basal spinules and outer element slightly longer than inner one.
Description of male. Based on allotype and four other paratypes. Body length from 279 to 312 Μm (mean = 302 µm, n = 5). Genital somite and third urosomite not fused ( Fig. 14 View FIGURE 14 D). Habitus ( Fig. 13 View FIGURE 13 B), colour, rostrum ( Fig. 13 View FIGURE 13 C), shape and ornamentation of cephalothorax and free prosomites ( Fig. 13 View FIGURE 13 B), shape and ornamentation of last three urosomites ( Figs. 13 View FIGURE 13 B, 14D), general shape, armature and ornamentation of caudal rami ( Figs. 13 View FIGURE 13 B, 14D), antenna ( Fig. 13 View FIGURE 13 B), labrum ( Fig. 13 View FIGURE 13 H), paragnaths ( Fig. 13 View FIGURE 13 H), mandibula ( Fig. 13 View FIGURE 13 H), maxillula ( Fig. 13 View FIGURE 13 H), maxilla ( Fig. 13 View FIGURE 13 H), maxilliped ( Fig. 13 View FIGURE 13 H), and all swimming leg ( Figs. 13 View FIGURE 13 B, 15J) as in female. Prosome/urosome ratio about 1.1, greatest width at posterior end of cephalothorax, body length/width ratio about 3.5; cephalothorax 1.6 times as wide as genital somite in dorsal view.
Genital somite ( Fig. 14 View FIGURE 14 D) 1.5 times as wide as long in ventral view, as in E. apimelon , but with more pronounced central internal ridge and fewer spinules at base of reduced sixth legs; single large spermatophore ( Fig. 15 View FIGURE 15 M) visible inside genital somite, about 3.2 times as long as wide without its neck.
Third urosomite ( Figs. 13 View FIGURE 13 B, 14D) as posterior part of genital double-somite in female but proportionately narrower, and with one or two large ventro-lateral spinules.
Caudal rami ( Figs. 13 View FIGURE 13 B, 14D) very similar to those in female but longer in proportion to anal somite and generally more elongated.
Antennula ( Figs. 13 View FIGURE 13 D, E, F, G, 14E, F) as in E. apimelon , except fourth segment with two delicate parallel combs ( Fig. 13 View FIGURE 13 E) instead of brush, additional small seta (or tubular pore?) present on fifth segment ( Fig. 13 View FIGURE 13 G) and long apical seta on that segment swollen at base, fourth segment slightly larger.
Fifth leg ( Fig. 15 View FIGURE 15 K, L) somewhat smaller than in female but with similar basic structure; exopod without proximal lateral armature element and with shorter spinules. Endopodal lobe without distal armature element, with relatively larger inner spines, and with smaller spinules on its three spiniform processes; endopodal inner spines of similar length and width.
Sixth legs ( Fig. 14 View FIGURE 14 D) as in E. apimelon ,simple cuticular plates, unarmed, with transverse row of several slender spinules.
Variability. Despite numerous examined specimens and detailed examination using SEM (see Figs. 10 View FIGURE 10 A, G, 11A, C, 12A, C, E, 13B), we are not able to report on any significant intraspecific morphological variability. The number of ventro-lateral spinules on the third urosomite in male varies between zero and three.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |