
Elaphinis (Micrelaphinis) pumila Boheman, 1857
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.004 |
|
publication LSID |
lsid:zoobank.org:pub:0F0B8FD8-76B1-44E0-BEDF-77628E7079EA |
|
persistent identifier |
https://treatment.plazi.org/id/73219E23-FFAF-FF85-1456-FE53FBB9FE6C |
|
treatment provided by |
Felipe |
|
scientific name |
Elaphinis (Micrelaphinis) pumila Boheman, 1857 |
| status |
|
Elaphinis (Micrelaphinis) pumila Boheman, 1857 ( Figs 24–34 View Figs 24–34 )
Elaphinis pumila Boheman, 1857: 40 .
Micrelaphinis pumila : PඣඋංඇGඎൾඒ (1907): 340, 341; SർHൾඇKඅංඇG (1921): 306; KඋൺඃčíK (1998): 53; BൾංඇHඎඇൽඇൾඋ (2017): 93.
Heteroclita (?) scitula Janson, 1878: 300 ; PඣඋංඇGඎൾඒ (1908): 684; SർHൾඇKඅංඇG (1921): 81; Mൺඋൺංඌ & Hඈඅආ (1989): 29.
Micrelaphinis (?) scitula : SർHൾංඇ (1960): 100.
Type material. Elaphinis pumila : Lൾർඍඈඍඒඉൾ: J (designated by Mൺඋൺංඌ & Hඈඅආ 1989), ʻCaffrariaʼ, ʻJ. Wahlb.ʼ, ʻType, pumila Bhn. Lnd. Caffr. ʼ, ʻRiksmuseum Stockholmʼ, ʻ462/85ʼ, ʻ Micrelaphinis pumila Boheman , Lectotype Holm 1986ʼ (TMSA-CPH1980). Pൺඋൺඅൾർඍඈඍඒඉൾ: J, ʻCaffrariaʼ, ʻJ. Wahlb.ʼ, ʻ463 35ʼ, ʻ Micrelaphinis pumila Boheman (var. a), Paralectotype, Holm 86ʼ, ʻ5378 E92 +ʼ, ʻRiksmuseum Stockholmʼ, ʻ(NHRS-JLKB 000027205)ʼ.
Heteroclita (?) scitula : Lൾർඍඈඍඒඉൾ: J, designated by Mൺඋൺංඌ & Hඈඅආ (1989), ʻNatalʼ, ʻMuseum Leiden verz. F.T. Valck Lucassen (O. E. Janson)ʼ, ʻ Heteroclita scitula, O. Jans. , Type, S. Africaʼ, Elaphinis (Micrelaphinis) pumila Boheman , det. E. Marais 1987ʼ, ʻ Heteroclita scitula Janson , Lectotypeʼ, ʻ( RMNH.INS.1266743)ʼ.
Additional material examined. 1 J, 85/28, corpulenta Gerst., Heteroclita (?) sp. bei scitula , det. H. Schein 1955, pumila (TMSA-CPH1981) ; 1 J, Natal, Durban, 11.1900, M. pumila Boh. , comp LT; PLT (via): STO, ʻCaffraria, J. Walhb.ʼ, M. scitula Jans. , comp. T: LEIDEN, ʻNatalʼ (TMSA-CPH1982).
Remarks. The status of this species remains rather enigmatic even in the present day and, therefore, deserves a dedicated analysis. To start with, most specimens identified under this name in historical collections have actually been proven to be colour varieties of E. (M.) delagoensis with a brown livery similar to that exhibited by inland specimens of E. (M.) irrorata (cf. Mൺඋൺංඌ & Hඈඅආ 1989, BൾංඇHඎඇൽඇൾඋ 2017; pers. obs.). Thus, there are only five specimens, all collected in the 1800s or early 1900s, that can be attributed to this species with confidence. These include the lectotype (LT) and one paralectotype (PLT), both males, from the original collections undertaken by Wahlberg in the eastern part of South Africa (ʻCaffrariaʼ – see notes below) during the period 1838–1845. Although these type specimens were only designated by Mൺඋൺංඌ & Hඈඅආ (1989), it is almost certain that they represent indeed the material used by BඈHൾආൺඇ (1857) in his original description of the species. Two other specimens are currently reposited in the TMSA and show all the typical characters of the species, but have only negligible traces of white tomentum on their dorsum and probably represent the ʻmodelʼ used by Mൺඋൺංඌ & Hඈඅආ (1989) in their illustration of this species (p. 26, fig. 58), later also reproduced in Hඈඅආ & Mൺඋൺංඌ (1992: pl. 21, fig. 5). A dorsal view of the PLT specimen has already been included in Bൾංඇ- Hඎඇൽඇൾඋ (2017) and is also reproduced here, along with a complete set of images obtained from Johannes Bergsten of the Naturhistoriska riksmuseet, Stockholm (courtesy: G. Beinhundner) ( Figs 24–30 View Figs 24–34 ).
A fifth specimen, again a male, originally described with some reservation as ʻ Heteroclita (?) scitula ʼ by Jൺඇඌඈඇ (1878), was later questioned by PඣඋංඇGඎൾඒ (1908) in terms of its generic affiliation and eventually transferred to Micrelaphinis by SർHൾංඇ (1960). It was only in the later revision of the genus by Mൺඋൺංඌ & Hඈඅආ (1989), however, that this species was synonymized with Elaphinis (M.) pumila and the specimen designated as LT. This specimen is reposited in the Naturalis Biodiversity Center, Leiden (RMNH), and a set of photos highlighting its critical diagnostic characters was recently obtained by courtesy of Oscar Vorst. This is now reproduced here as Figs 31–34 View Figs 24–34 , in order to corroborate the validity of its earlier synonymy and highlight the remarkable variability in dorsal tomentose maculation exhibited by the species. Indeed, this can range from virtual complete absence, with only residual spots or lining on the pronotal margins (cf. Hඈඅආ & Mൺඋൺංඌ 1992: pl. 21, fig. 5), to a rather dense and intricate network of round and irregularly shaped maculae and stripes, both on the dorsal and ventral surfaces ( Figs 31–33 View Figs 24–34 ). Consistent features across the range are, however, its dense pubescence, which is particularly noticeable on head and pronotum, and the testaceous background colour of its elytra. These, along with the uniquely shaped apical expansions of its aedeagal parameres, make it possible to separate it fairly promptly from its closest relatives, i.e. E. (M.) latecostata and E. (M.) delagoensis (cf. identification key below).
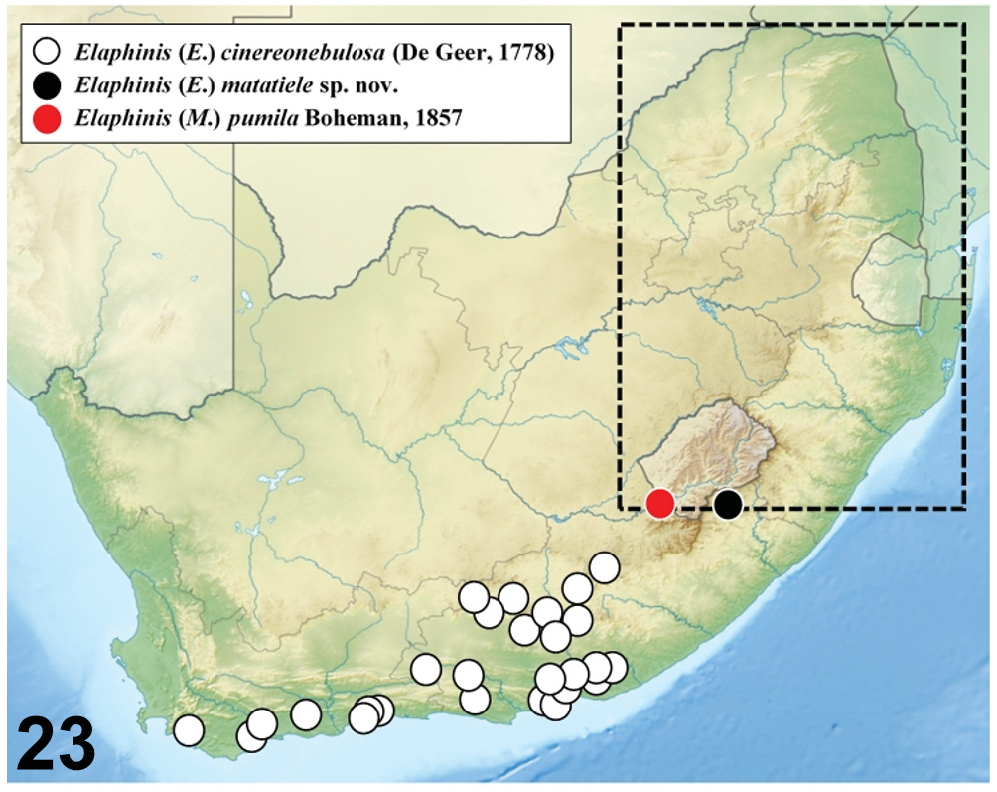
Regarding the correct geographic origin of the two type specimens collected by Wahlberg, BඈHൾආൺඇ (1848) explained in the introductory section that Wahlberg’s ʻCaffrariaʼ included the north-eastern part of South Africa, from approximately 30°25′ to 23°00′S and 32°40′ to 27°00′E (cf. black dotted frame within map in Fig. 23 View Fig ). This includes an area comprising virtually the entire provinces of KwaZulu-Natal, Mpumalanga, Limpopo and Gauteng, along with the eastern half of the Free State, the extreme eastern portion of the North-West and only the north-eastern tip of the Eastern Cape Province, to the north of the town of Lady Grey and bordering Lesotho. This is drastically different to the historical definition of ʻKaffrariaʼ used by the British, who regarded this region as comprising the entire eastern half of what is today the Eastern Cape Province (CඋංඌHඈඅආ 1911). Regarding the type locality of E. (M.) pumila , BඈHൾආൺඇ (1857: 40), specifically stated ʻ Habitat in regione fluvii Gariepis ʼ. As this unequivocally refers to the Orange River valley, the only possibility within the area sampled by Wahlberg in South Africa lies in the south-eastern part of the Free State, south of Zastron, or in the north-eastern tip of the Eastern Cape, north of Lady Grey. Alternatively, this may also be located in neighbouring Lesotho. The distribution map illustrated in Mൺඋൺංඌ & Hඈඅආ (1989: 24, fig. 48), shows only one locality ʻNatal, Durban (SE 2931 Cc)ʼ, while the revised map presented in Hඈඅආ & Mൺඋൺංඌ (1992: 181, fig. 98g) has an additional locality in the border area between KwaZulu-Natal and Mpumalanga, near the town of Wakkerstroom. It is not clear how these localities were derived, as with the exception of the type specimens there is no specific list of specimens label data in either of the two works mentioned above. The locality ʻNatal, Durbanʼ is reported on one label of specimen CPH1982 (TMSA), but this locality is almost certainly erroneous and probably reflects the ʻport-of-callʼ role that this city used to play in the early days of entomological collections in the eastern part of South Africa. A series of four specimens carrying the identification of ʻ M. pumila Boh. ʼ reposited in the ZMHB also has the locality ʻNatal/Durbanʼ in their accompanying labels, but they are actually all males of E. (M.) delagoensis , lacking in particular the dense and long pubescence characteristic of Boheman’s E. (M.) pumila LT and PLT ( Figs 24–26 View Figs 24–34 ).
What emerges rather clearly from this review is that E. (M.) pumila is not coastal, but rather a highland dweller possibly restricted to a small region above the Drakensberg Escarpment. As already pointed out by Mൺඋൺංඌ & Hඈඅආ (1989) and Hඈඅආ & Mൺඋൺංඌ (1992), what is remarkable though is that no new specimens are known besides the types collected in the 19–20 th centuries. So, it is possible that the species may indeed be already extinct, as extensive land-use changes have happened in its hypothetical distribution range, both due to agricultural activities and human settlements.

| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Elaphinis (Micrelaphinis) pumila Boheman, 1857
| Perissinotto, Renzo 2022 |
Heteroclita
| JANSON O. E. 1878: 300 |
Elaphinis pumila
| BOHEMAN C. H. 1857: 40 |