
Elaphinis (Elaphinis) matatiele, Perissinotto, 2022
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.004 |
|
publication LSID |
lsid:zoobank.org:pub:0F0B8FD8-76B1-44E0-BEDF-77628E7079EA |
|
persistent identifier |
https://treatment.plazi.org/id/73219E23-FFAA-FF89-17E2-FCA1FA3EFCEC |
|
treatment provided by |
Felipe |
|
scientific name |
Elaphinis (Elaphinis) matatiele |
| status |
sp. nov. |
Elaphinis (Elaphinis) matatiele sp. nov.
( Figs 1–9 View Figs 1–9 , 19 View Figs 19–22. 19–20 )
Type material. Hඈඅඈඍඒඉൾ: J, South Africa, EC, Matatiele, 7 Dec 2008, R Perissinotto & L Clennell ( ISAM). Pൺඋൺඍඒඉൾඌ: 11 JJ 1 ♀, same data ( BMCS, RPGS); 1 J 1 ♀, RSA, Eastern Cape, Alfred Nzo District (Matatiele Municipality), Matatiele Nature Reserve, 1900 - 2100 m, 1 Jan 2017, P. Malec & P. Šípek leg. ( PMBC, UKPC); 1 J, same locality, Bred from wild larvae, P. Malec breeding ( PMBC); 58 JJ 3 ♀♀, same locality, 6 Dec 2019, P. Šípek, M. Hiřman, O. Kouklík leg. ( NMPC, OKPC, PMBC, UKPC); 3 ♀♀, same locality, Dec 2019, P. Šípek leg., ex ovum, P. Malec Breed F1 ( PMBC).
Description of holotype male ( Figs 1–8 View Figs 1–9 ). Body. Black and matte, with head, pronotum and scutellum predominantly black but elytra extensively covered in white-greyish (often discolouring to pink or even brick-red in preserved specimens) tomentum, except on periscutellar, perisutural and postero-lateral regions; with regularly spaced round sculpture on head and apical part of pronotum (around tubercular ridge), becoming minute and very scattered on the pronotal disc, scutellum and elytra; with dense and long, pale-yellow to tawny setae on entire head surface, becoming shorter, more scale-like in shape and scattered on pronotal declivities and mesepimera, and virtually disappearing on pronotal disc, elytra and scutellum ( Figs 1–4 View Figs 1–9 ).
Size. Total length = 11.9 mm; maximum width = 6.1 mm.
Head. Entirely black and concave; clypeus broadly bilobate, with angulate indentation at apex and lateral margins smoothly rounded; all margins sharply upturned, especially in apical region around indentation; entire surface covered in round to coarsely irregular sculpture; pale-yellow to tawny setae distributed along entire surface, generally shorter and more scattered on clypeus and frons, becoming much longer and denser on vertex; antenna dark brown to black, with club approximately as long as flagellum and pedicel combined; thin light setae scattered across flagellum, becoming thicker and denser on pedicel ( Fig. 4 View Figs 1–9 ).
Pronotum. Black and matte, with white-greyish tomentose ornamentation along lateral margins spreading towards declivities on posterior half and generating two short converging lines in suprascutellar area; with tubercular ridge in central apical area exhibiting dense round punctures on declivities; punctures becoming smaller and more scattered along lateral margins and declivities but minute on disc; short scale-like setae distributed along lateral margins and declivities, becoming minute on disc; shape smoothly rounded along lateral and posterior margins, with very weak obtuse angle above scutellum; antero-lateral margins abruptly angulate, leading to steep declivity up the apical tubercle ( Figs 1, 3 View Figs 1–9 ).
Scutellum. Black, matte and glabrous, with tomentose lining around lateral and apical margins; few, shallow round to crescent or irregularly shaped punctures along basal and part of lateral margins, inwards of tomentose lining; broadly isoscelic triangular in shape with smoothly round apex; lateral grooves very narrow and shallow, deepening somehow around apex ( Fig. 1 View Figs 1–9 ).
Elytron. Black and matte, with shiny patches in umbonal areas; with intricate white-greyish (often discolouring to pink or even brick-red in preserved specimens) ornamentation of maculae and dotted stripes across entire surface, excluding umbonal, periscutellar, perisutural and posterolateral areas; black areas on elytral disc producing symmetric figure in shape of inverse anchor when both elytra are combined; costae obsolete to weakly elevated and barely visible; striae equally faded, exhibiting some shallow and scattered round punctures; humeral callus protruding remarkably outwards, leading to formation of deep subhumeral arch; posterior margin smoothly rounded without projections or upturning at apex; virtually glabrous on disc and umbones, but with scattered short and light setae distributed along lateral and apical declivities ( Figs 1, 3 View Figs 1–9 ).
Pygidium. Broadly triangular in shape, with smoothly rounded apex and unevenly domed with marked depressions on baso-lateral corners; exhibiting white-greyish tomentose markings most noticeable in basal and lateral regions; with dense but shallow rugulose sculpture across entire surface and light-yellow to tawny short setae scattered throughout disc, becoming longer and finer along lateral margins and apex ( Fig. 5 View Figs 1–9 ).
Legs. Black and shiny with occasional brown tips or edges; tarsomeres rather elongate, with apical ones twice as long as preceding units and all claws distinctively bent at 90° angles; tibiae irregularly sculptured and covered in dense, scale-shaped, light-yellow setae, becoming longer and finer on inner margins; protibia broad and bidentate, with both teeth equally developed and residual presence of regressive third tooth in proximal area; mesotibia with bifid mid spine on outer carina sharply pointed and spurs elongate, thin and sharp; metatibia with outer carina bearing one blunt but robust distal tooth, spurs substantially more elongate than in mesotibia and remarkably sharp ( Figs 1–3 View Figs 1–9 ).
Ventral surface. Black and shiny, with white-greyish tomentum markings restricted to marginal areas of ventrites; covered in dense and long light-yellow to tawny setae, except on central metafemoral surface, abdominal ventrites and mesometasternal region, where setae become short or very scattered; mesosternal lobe very narrow, smoothly rounded and not protruding either forward or downwards; metasternal lobe exhibiting oblong, but very narrow and subparallel groove on anterior two thirds of median sulcus; abdominal ventrites with slight concavity at centre ( Fig. 2 View Figs 1–9 ). Aedeagus. Parameres with dorsal lobes relatively elongate and exhibiting a narrow constriction both towards base and apex, before expanding again in both directions ( Figs 6–8 View Figs 1–9 ); apical projection of lobes curving smoothly downwards and forming regularly rounded, drop-shaped tips in frontal view ( Fig. 8 View Figs 1–9 ); ventral lobes wider than dorsal virtually across entire length, reaching maximal expansion at mid to basal distance and forming a small pair of lateral projections in dorsal and frontal views ( Figs 6, 8 View Figs 1–9 ).
Variability. Specimen size lies in the range of 10.9–13.1 mm in total length and 6.1–6.7 mm in maximum width. The dorsal ornamentation can vary in extent on the pronotum, elytra and pygidium, but its general pattern remains rather stable across different individuals. Females are stockier than males, have broader protibia, shorter antennal clubs and protarsi, exhibit reduced tomentose ornamentation on the dorsum and complete absence on the venter ( Fig. 9 View Figs 1–9 ). They are also characterized by drastically reduced pubescence, by comparison with their male counterparts, and therefore tend to appear shiny. Their dorsal surface is, however, more densely covered in round punctures, particularly on head, pronotum and protibiae.
Differential diagnosis. This is clearly a sister species of Elaphinis cinereonebulosa , with which it shares all the key characters of the nominal subgenus. The two species can easily be separated even on the basis of their dorsal and ventral ornamentation, as the tomentose markings are extensive in E. cinereonebulosa but substantially reduced in E. matatiele sp. nov., particularly on the pronotal, scutellar and elytral discs as well as abdominal ventrites ( Figs 1, 3 View Figs 1–9 ). Other notable diagnostic differences are observed at the level of the clypeus, the scale-shaped setae of the general body cover and the aeadeagal parameres ( Figs 1–8 View Figs 1–9 ). The clypeus is slightly longer and narrower in E. matatiele sp. nov., by comparison with that of E. cinereonebulosa , and also exhibits a sharp angular indentation at the apex in the former, rather than a smooth sinuation like in the latter species ( Figs 4 View Figs 1–9 , 13 View Figs 10–18 ). The scale-shaped setae that cover much of the dorsal surface of E. cinereonebulosa are remarkably shorter and broader than those of E. matatiele sp. nov. Also, they are very dense in many areas (e.g. head and antero- -pronotal surface) on the dorsum of E. cinereonebulosa , while they are substantially more scattered in all the areas of E. matatiele sp. nov. Finally, the aedeagal parameres of E. matatiele sp. nov. are substantially narrower and slightly longer than those of E. cinereonebulosa ( Figs 6–8 View Figs 1–9 , 15–17 View Figs 10–18 ). The ventral lobes of the parameres are particularly well developed in E. cinereonebulosa , with baso-lateral projections much larger in extent than those observed in E. matatiele sp. nov. ( Figs 6 View Figs 1–9 , 15 View Figs 10–18 ).
Etymology. The species is named after its type locality, which is also the only place where it is currently known to occur. The specific epithet is a noun in the nominative singular.
Bionomy. Adult activity in the wild has so far been recorded only in December and January. All specimens were retrieved from sandy soil accumulations in the proximity of, or under, old cow dung on rocky terraces at high altitude ( Fig. 19 View Figs 19–22. 19–20 ). Larvae of the species bred in captivity have shown very long pre-pupal dormancy and high mortality rates, thereby proving it difficult to reach adult stage under controlled conditions (P. Malec, pers. comm.).
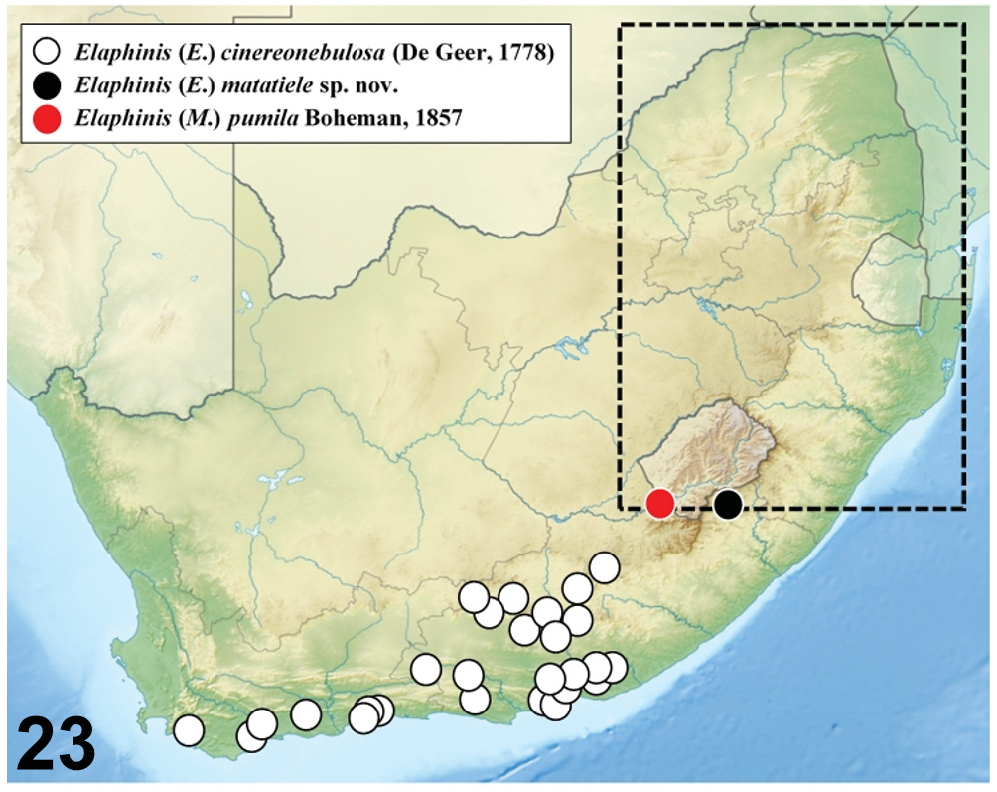
Distribution. All the specimens currently known originate from the Matatiele Nature Reserve ( Fig. 23 View Fig ), which is a core protected area within the Maloti-Drakensberg Transfrontier Conservation Area. The Matatiele Municipality was part of Cape Province until being transferred to Natal in 1978; in 2005 it was moved back from the KwaZulu-Natal Province to the Eastern Cape. Most specimens of the new species were collected around the base of Mount Hargreaves, at an altitude of about 1700 m ( Fig. 21 View Figs 19–22. 19–20 ), and it is virtually certain that it also occurs on several nearby mountains that have similar habitat characteristics. These, however, have not been explored recently due to issues of accessibility.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.