
Eclipidrilus pacificus, Steven & Fend, V., 2005
|
publication ID |
https://doi.org/ 10.5281/zenodo.171279 |
|
DOI |
https://doi.org/10.5281/zenodo.6266040 |
|
persistent identifier |
https://treatment.plazi.org/id/039F8792-6D01-F143-FE8E-FD5AA689F964 |
|
treatment provided by |
Plazi |
|
scientific name |
Eclipidrilus pacificus |
| status |
sp. nov. |
Eclipidrilus pacificus n. sp. ( Figures 1–10 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
HOLOTYPE: USNM 1073033. A whole mounted worm in Kleermount.
TYPE LOCALITY: Washington: Jefferson Co.: pool in small spring next to the Hoh River, 47º49’00”N, 124º06’46”W, in mud and macrophyte roots in shallow water.
PARATYPES: From the type locality. USNM 1073034–1073037: 2 sectioned worms (1 sagittal, 1 transverse), 1 whole mount, 2 dissected and slide mounted. Collected by S. Fend. 29.IV.1999.
OTHER MATERIAL: Typical specimens: Washington: Clallam Co: muddy seep beside Road 29, Calawah River drainage, 47º57’29”N, 124°14'31"W, 26.IV.2004. 4 dissected. Jefferson Co: from the type locality, 29.IV.1999. 1 whole mount, 1 longitudinally dissected. 2.VI.2003. 4 whole mounts, 4 dissected; 2 partially mature, dissected. Small tributary to Clearwater River at Coppermine Bottom Camp, 47°39'21"N, 124°11'58"W, 25.VI.2004. 8 dissected. Pacific Co.: South Naselle River near Knappton, 46°17'38"N, 123°48'36"W, 25.VI.2004. 5 dissected. All collected by S. Fend.
Variant 1: Idaho: Latah Co.: Palouse River at Laird Park, 46º56’35”N, 116º38’29”W, 2.V.2000, collected by D. L. Gustafson. 1 dissected. Shoshone Co.: Middle Fork Saint Maries River near Two Bit Creek, 47°00’08”N, 116°10’30”W, 28.VII.2003, collected by D. L. Gustafson. 2 dissected. Oregon: Yamhill Co.: small spring west of Peavine Ridge, in mud and macrophyte roots, slow current, 45º14’19”N, 123º 22’50”W, 30.I.2000. 1 whole mount, 1 dissected, 2 sectioned (1 sagittal, 1 transverse). 10.V.2001. 6 whole mounts. Lane Co.: small spring and stream beside Fox Hollow Rd., Eugene, in mud, 44º00’36”N, 123º04’50”W, 29.I.2000. 3 whole mounts. 1.VI.2003. 4 dissected, 3 whole mounts, 2 sectioned (1 sagittal, 1 transverse). Spring at mouth of Tenmile Creek, in mud and macrophyte roots, shallow water with slow current, 44º13’30”N, 124º06’29”W, 30.I.2000. 2 whole mounts. Washington: Pierce Co.: Taylor Bay Inlet, 9.IX.2003, collected by M. Allen. 3 dissected. Worms collected by S. Fend unless otherwise noted.
Variant 2: Oregon: Clatsop Co.: Columbia River estuary at Snag Island, VIII.1980, collected by E.J. Clairain and C. Newling. 8 whole mounts, from the Royal Ontario Museum, No.1989037, slides labeled 1, 5, 7, 8, 9, 11, 16, 17. Karlson Island ( Columbia River estuary), 28.VI.2002, collected by R. Draheim. 1 whole mount, 3 dissected, 2 sagittally sectioned. Knappa Slough at dock ( Columbia River estuary), 24.VI.2002, collected by R. Draheim. 5 dissected. Warren Slough ( Columbia River estuary), 28.VI.2002, collected by R. Draheim. 2 whole mounts, 1 dissected. Tillamook Co.: spring beside Nestucca River, 7 km upstream of Blaine, 4.VI.2003, collected by S. Fend. 1 dissected.
Partially mature, undetermined: Pacific Co.: small spring along the Naselle River, 46º24’00”N, 123º43’42”W, in mud and macrophyte roots, 30.IV.1999, collected by S. Fend. 1 dissected. Pierce Co.: Little Mashell Creek, 20.IX.2001, collected by M. Allen. 1 whole mount.
Description
Typical specimens
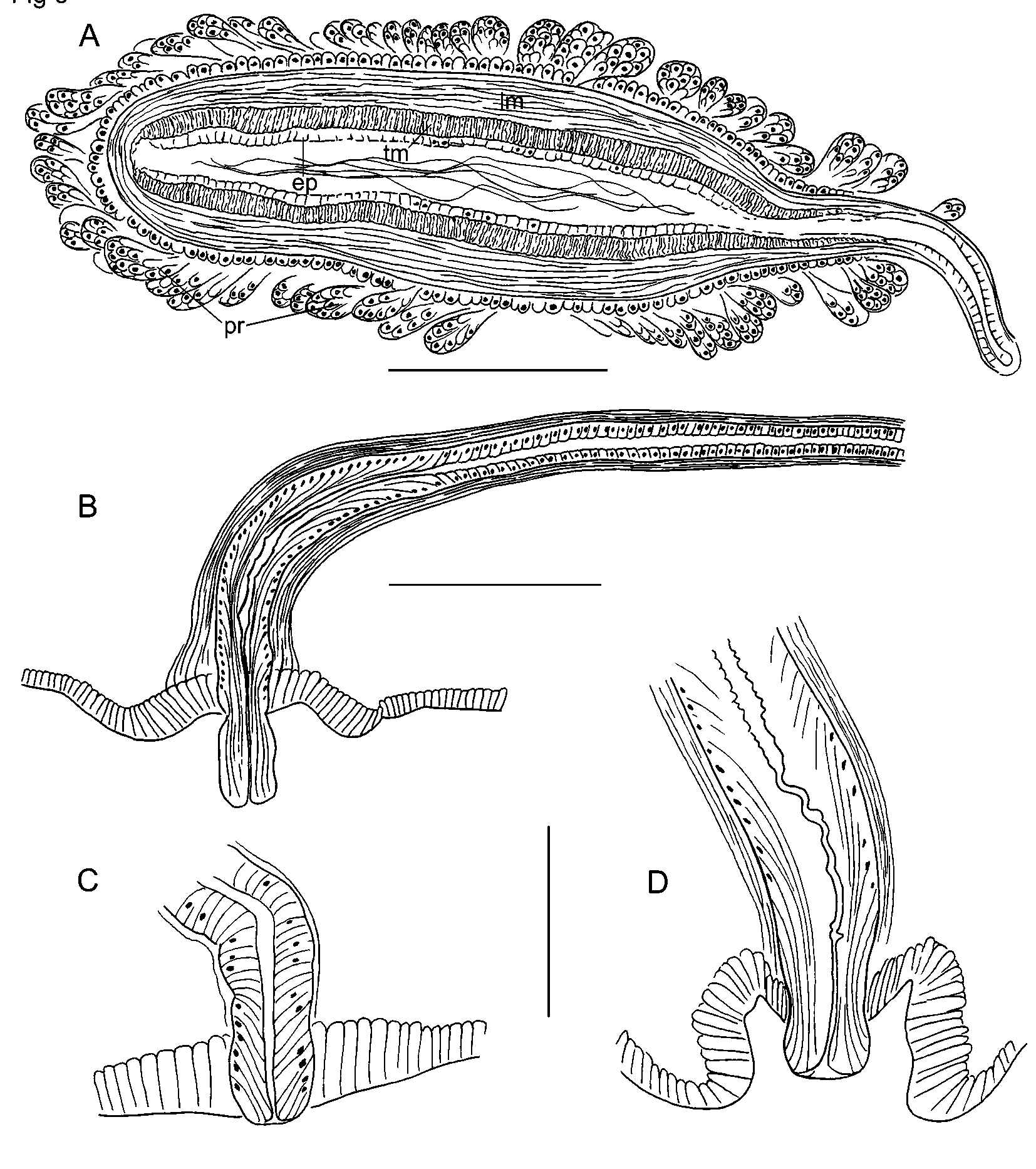
Length of preserved worms 17–32 mm, 69–107 segments; width 0.52–0.80 mm in X, maximum width to 0.82 mm. Secondary segmentation prominent in segments V–X, resulting in a narrow anterior ring in each segment; variable in posterior segments ( Fig. 1 View FIGURE 1 A–B). Chaetae sigmoid, simplepointed, with nodulus 29–38% of chaeta length from tip ( Fig. 2 View FIGURE 2 A). Chaeta length 125–170 µm in midbody; slightly shorter in posterior segments, but proportions similar in anterior and posterior segments. Dorsal and ventral chaetae approximately equal in length; within each bundle, the outer (more lateral) chaeta slightly shorter than the inner. Prostomium roundedconical, about 0.3 mm long, width about equal to length.
Brain in I, not strongly lobed. Pharynx in II–IV; dorsal wall with columnar cells; ventral wall much thinner than dorsal anterior to IV ( Fig. 2 View FIGURE 2 B, cf. Fig. 14 View FIGURE 14 A). Pharyngeal glands in V–VII(VIII). Longitudinal muscle layer 20–35 µm thick in preclitellar segments; circular muscle layer 5–7 µm; both layers thinner in posterior segments. Epidermis 10–20 µm thick anterior to clitellum; to about 35 µm in clitellum.
Dorsal blood vessel passes under the brain in I before dividing; the two forks pass around the buccal cavity and rejoin in V as a single ventral vessel. One pair of lateral, commissural blood vessels joins dorsal vessel in segments anterior to XII; each commissure strongly convoluted, and those in the first few segments usually enter the next segment before joining the ventral vessel ( Fig. 2 View FIGURE 2 B). Commissures in X and XI loop into sperm and egg sacs. Dorsal vessel without lateral vessels between the clitellar segments and about the posterior 1/2 to 1/3 of the body. Beginning near midbody or posteriorly, two pairs of short, blind lateral blood vessels usually branch off the dorsal vessel in each segment. Lateral vessels are longer, with up to 5–10 short branches in the posterior 1/3 to 1/4 of some specimens ( Fig. 2 View FIGURE 2 D), but remain short and inconspicuous in others ( Fig. 2 View FIGURE 2 C). Perivisceral sinus begins in VII; chloragogen cells begin in VII or VIII.
First nephridia usually paired on 6/7, the next on 12/13 or posterior; nephridia in few to most posterior segments, paired or on one side only. Each nephridium has a small anteseptal funnel, a granular postseptal thickening, and an irregular, posterior duct 18–20 µm in diameter. The posterior duct branches, with a short branch to the nephropore anterior to ventral chaetae. Another branch extends along the ventral blood vessel as a convoluted mass, entering posterior and/or anterior segments; posterior to XIII it appears continuous with ducts from other nephridia across multiple segments. Nephropores simple, inconspicuous.
Testes paired in IX and X, smaller in IX. Ovaries paired in XI; usually elongate, extending to 11/12 or into XII. Sperm sacs paired, extending back as far as XXII; egg sacs may extend 1 or 2 segments beyond sperm sacs. Female funnels up to 120 µm tall, with the posterior side much longer than the anterior; female pore intersegmental, on 11/12. Clitellum IX–XIII.
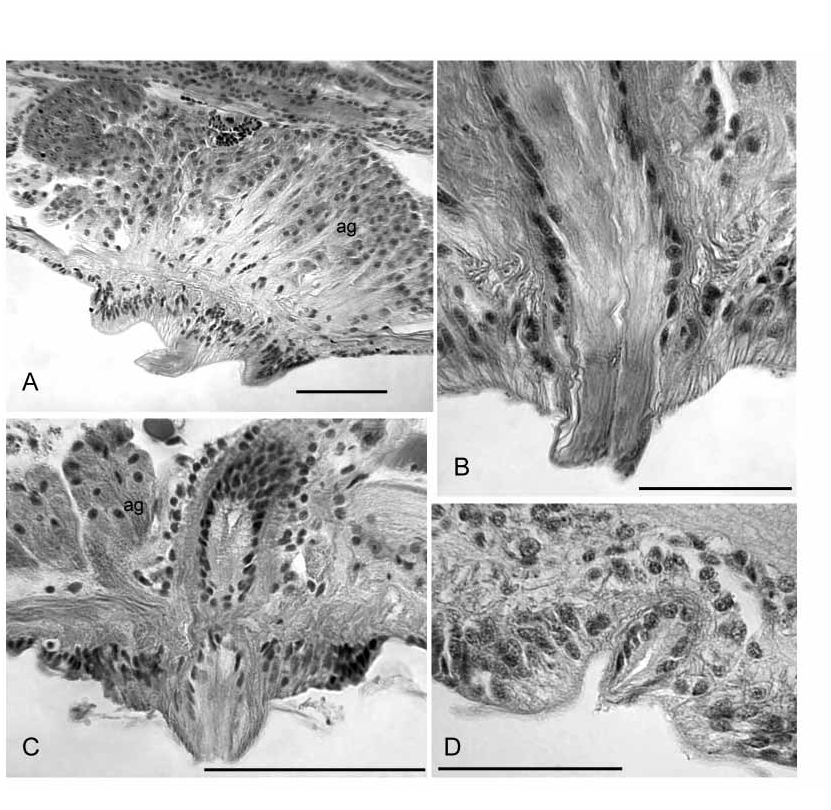
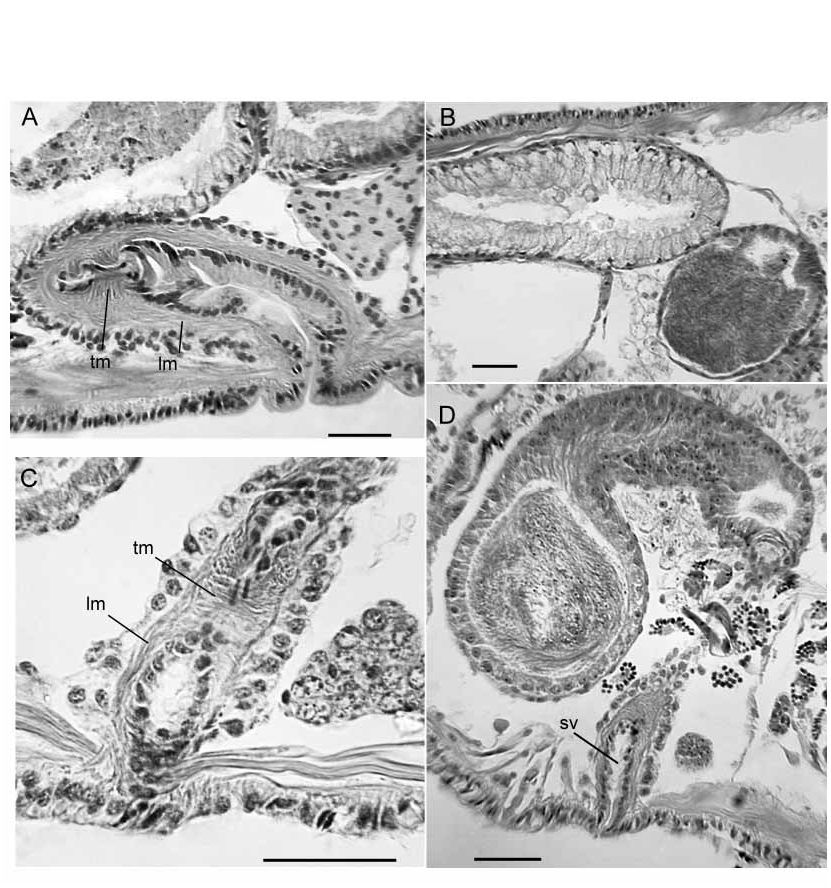
Single male pore midventral (beneath the ventral nerve cord) in X, behind the ventral chaetae ( Figs. 1 View FIGURE 1 A–B, 3A, 4A). A short, cylindrical penis is contained within a shallow concavity, within a ring of thickened epidermis 170–340 µm in diameter ( Figs. 4 View FIGURE 4 A, 5B,D, 8A–B). Epidermal cells within the concavity interspersed with narrow ducts from large (160–300 µm tall), petiolate accessory glands; gland mass to 550 µm wide ( Figs. 4 View FIGURE 4 A, 8A). Penis variably extended, 20–160 µm long and 40–50 µm wide, formed by extrusion of the atrial duct epithelium. Epithelial cells in the most ectal part of the duct have basal nuclei and fibrous, ectallydirected extensions that appear continuous with the penis ( Figs. 4 View FIGURE 4 A, 5D, 8B). Lumen at ectal end appears slightly cuticular, and may appear irregular or sinuous ( Fig. 5 View FIGURE 5 D, 8B).
Atrium extending as far as XIX, with distinct ectal duct and ental ampulla ( Fig. 3 View FIGURE 3 A). Atrial duct 600–2300 (1530) µm long. The ectal half is a 60–100 µm diameter tube of loose, mostly longitudinal muscle, about 20 µm thick, with an inner epithelium that changes from columnar to elongatefibrous in about the ectal 200 µm, near the male pore ( Figs. 4 View FIGURE 4 A, 5B). Ental portion of duct about 40 µm in diameter with a thinner (4–6 µm) muscle layer that is mostly longitudinal, with a very thin (ca. 1 µm) inner circular layer.
Ampulla nearly cylindrical, but tapered ectally; length 520–1900 (890) µm, maximum diameter of muscle tube 130–220 (180) µm. Ampulla with an outer layer of longitudinal muscle 22–50 (38) µm thick, an inner transversecircular layer 10–25 (15) µm thick, and epithelium 8–19 µm thick ( Figs. 5 View FIGURE 5 A, 9A–C). Fibers in the longitudinal muscle layer in radially arranged lamellae ( Fig. 9 View FIGURE 9 B), parallel to the long axis of the ampulla, slightly sinuate, but not arranged in opposing layers. The inner, transverse muscle layer has parallel fibers arranged orthogonally to the long axis of the atrium ( Fig. 9 View FIGURE 9 A–C). Lumen of ampulla variable, 12–100 (42) µm wide, or 0.08–0.61 (0.27) times atrium diameter. The atrial ampulla is covered with a single layer of small cells (possibly nuclei of muscle cells) and a layer of densely packed, multicellular prostate glands ( Fig. 5 View FIGURE 5 A, 9A–C). Prostatic cells are narrowly petiolate, usually occurring in small bundles of up to about 10 cells; individual bundles 40–90 µm tall.
Male funnels paired on 9/10 and 10/11; the anterior pair smaller than the posterior (median height 100 µm vs. 160); the posterior pair usually extends back into XI within the sperm sac. Both anterior and posterior male funnels usually functional, with associated sperm. Both anterior and posterior vasa deferentia narrow (8–16 µm diameter). All four vasa deferentia join the atrial duct, running outside the muscle layer until the ampulla, where they gradually pass through the muscle layers ( Fig. 9 View FIGURE 9 B) and eventually the epithelium, and enter the ental part of the lumen. Posterior vasa deferentia enter X directly, without penetrating 10/11 and forming a loop in XI.
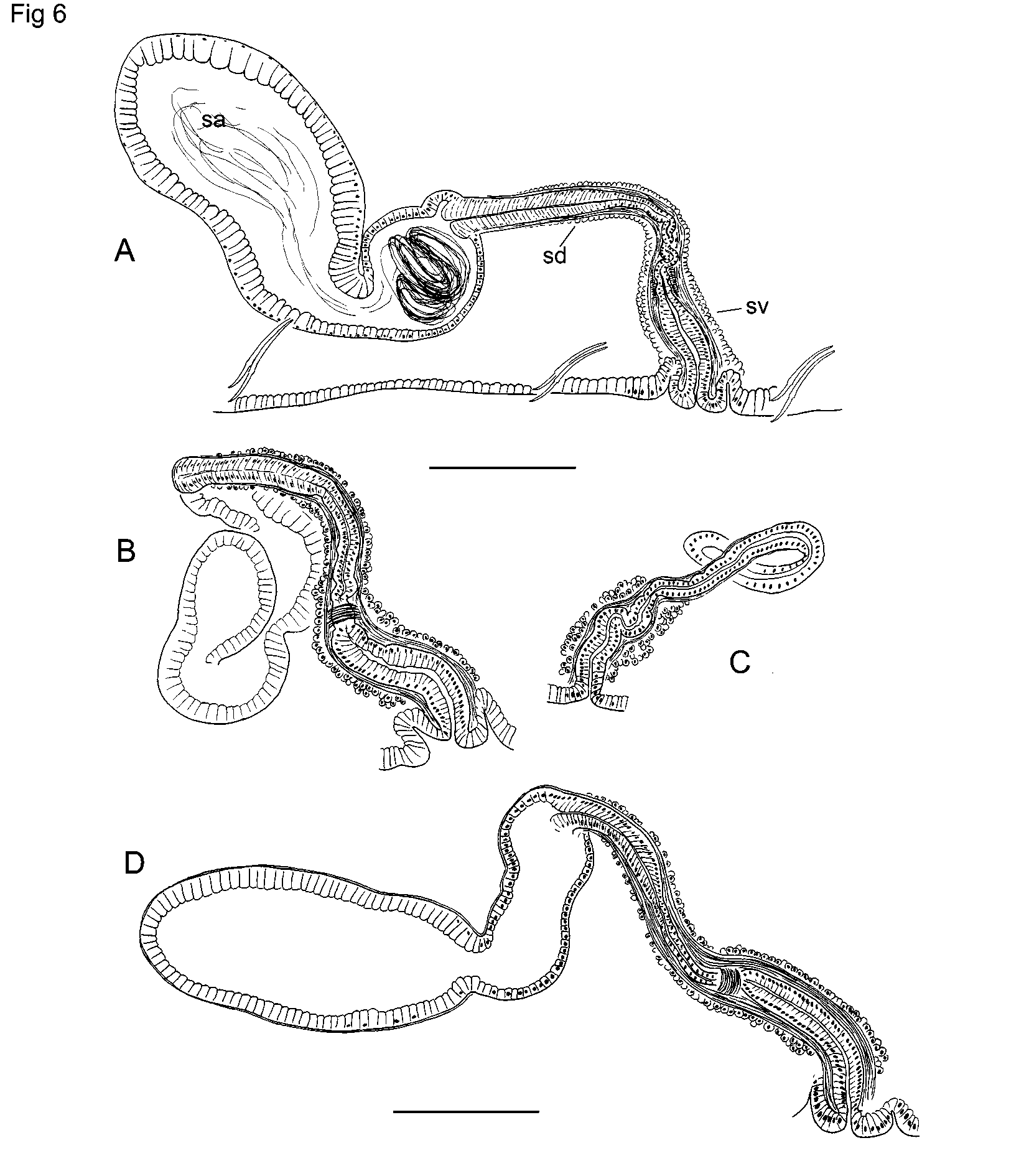
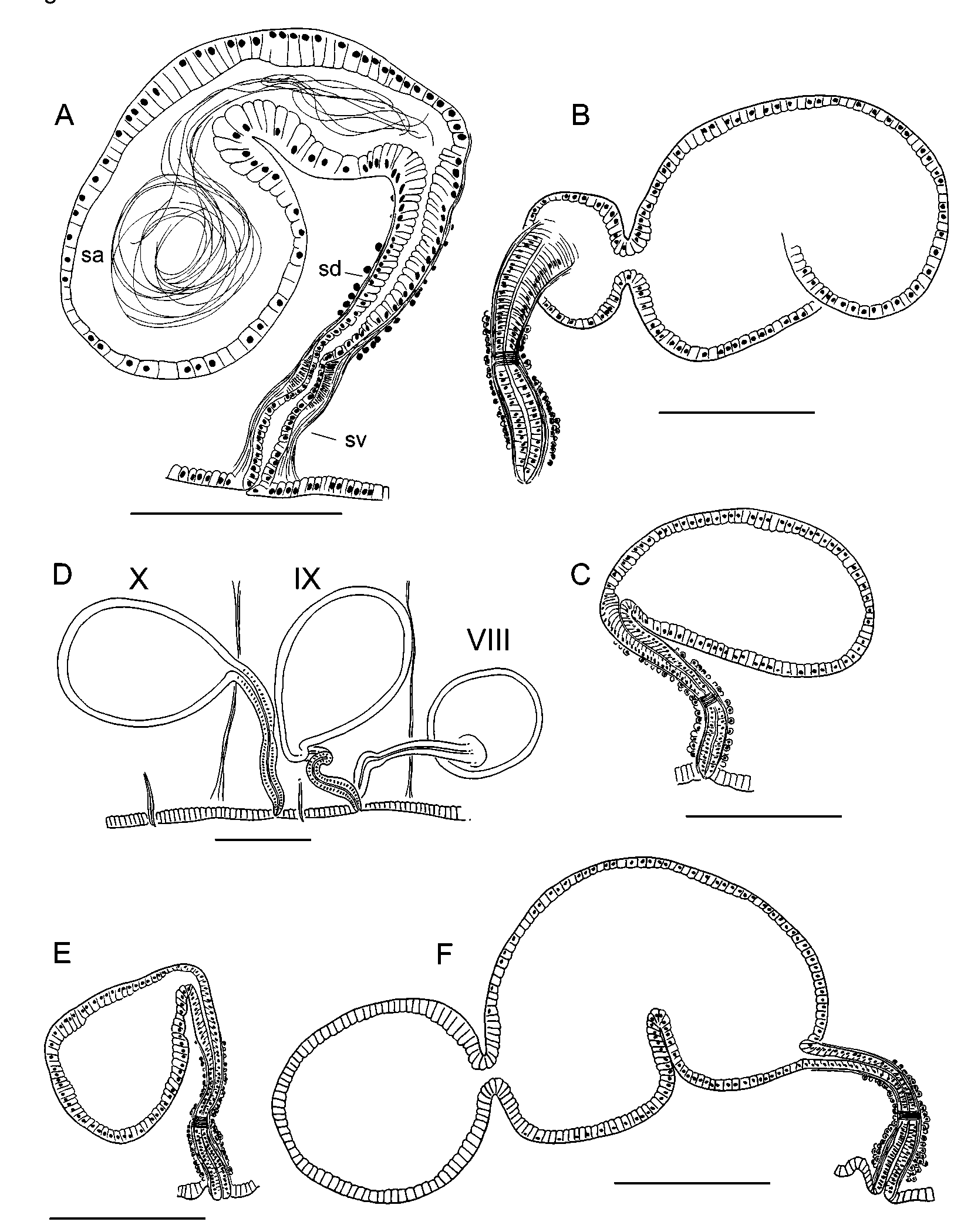
Spermathecae paired in IX; pores on ventral chaetal lines, near the anterior septum (8/ 9), usually within the narrow, secondary annulus ( Fig. 1 View FIGURE 1 A–B). Pores usually prominent, sometimes on low mounds. Each spermatheca has a narrow ectal duct 230–720 (370) µm long, and a distinct ental ampulla 420–1330 (680) µm long by 130–410 µm wide ( Fig. 6 View FIGURE 6 A,D). The ectal end of the duct widens into a narrow vestibule 145–310 (215) µm deep by 64–96 (77) µm wide, with a mostly longitudinal muscle layer surrounding a very thin transversecircular layer and columnar lining cells with basal nuclei ( Figs. 6 View FIGURE 6 A, 10A). Entally, the duct narrows abruptly, forming a short, convoluted section inside the muscle tube ( Fig. 6 View FIGURE 6 A). A short band of thickened, transversecircular muscle surrounds and constricts the beginning of this section ( Fig. 10 View FIGURE 10 A). Near the midpoint, the duct widens; the ental half is 40–50 µm in diameter; a thin (3–4 µm), muscle layer surrounds a columnar epithelium. Spermathecal ampullae in IX or IX and X, covered by a very thin muscle layer. Ampullar epithelium thin (4–5µm) in about the ectal 1/5, but entally becomes thicker (18– 42 µm) with columnar, granular cells having basal nuclei ( Figs. 6 View FIGURE 6 A, 10B). Cells may appear increasingly vacuolated, with very small nuclei, toward ental end; apparent sperm cells are visible within the vacuoles in some specimens. In most specimens the sperm is strongly concentrated near the duct ( Fig. 10 View FIGURE 10 B). The ental part of the ampulla usually contains only scattered sperm cells, and may contain an unknown, eosinstaining substance.
In partially mature worms the male pore is on a pad of slightly thickened epidermis, with welldeveloped accessory glands. Ectal end of duct with columnar (not fibrous) lining cells, protruding slightly as a small papilla ( Fig. 5 View FIGURE 5 C), similar to mature E. pacificus Va ri a nt 1 (see below). Atrial duct and ampulla weakly differentiated; muscle layers thin (less than 10 µm total), but inner epithelium of ampulla distinct, 5–15 µm thick. Atrial ampulla and duct covered by a dense layer of cells, which are not in clusters. Spermathecal ampulla not strongly differentiated from duct in early stages ( Fig. 6 View FIGURE 6 C), but becomes distinct prior to mating ( Fig. 6 View FIGURE 6 B). Ectal vestibule of spermatheca distinct, even at an early development stage ( Fig. 6 View FIGURE 6 C).
Variant 1
Length of preserved worms 12–26 mm, 53–86 segments; width 0.38–0.65 mm in X. Chaeta length in clitellar region 101–136 µm, nodulus 28–36 % of chaeta length from tip. Blood vessels sometimes as described for typical E. pacificus , but in many specimens the lateral blood vessels of posterior segments are very short and appear unbranched; in some specimens posterior lateral vessels are difficult to see, and may be absent.
Male pore less prominent than in typical E. pacificus ( Fig. 1 View FIGURE 1 C); usually surrounded by a concave pad of slightly thickened epidermis 75–180 µm wide, within a patch of nonclitellar epidermis. Atrial duct terminates on a small, conical papilla, which appears to be a simple continuation of the duct ( Fig. 8 View FIGURE 8 D), without extremely elongated or extruded lining cells.
Atrium usually extending to XII–XVI, but sometimes entirely in X. Atrial duct 330– 1870 (585) µm long; ampulla length 300–1480 (455) µm, diameter 84–194 (112) µm. Musculature of atrial ampulla variable, but usually much thinner than in typical specimens: longitudinal muscle 6–22 (8) µm thick; inner, transversecircular muscle 3–12 (6) µm ( Fig. 9 View FIGURE 9 D). Longitudinal muscle fibers parallel to ampulla or slightly spiral, with angle less than 20º from longitudinal axis. Epithelium of ampulla very thin, usually less than 3 µm; lumen 0.3–0.8 (0.64) times total ampulla width. Prostate glands to 25–50 µm tall. Both pairs of vasa deferentia very thin, diameter 6–12 µm. Anterior male funnels 60–110 µm wide, cupshaped; with or without sperm. Posterior funnels 100–200 µm wide, usually convoluted, and entering sperm sac to about the middle of XI.
Spermathecal pores inconspicuous; anterior to ventral chaetae, but position variable, usually posterior to the anterior, secondary annulus in IX ( Fig. 1 View FIGURE 1 C). Ectal vestibule of spermathecal duct with thinner musculature than in typical specimens; vestibule usually distinct, 55–150 µm long by 30–44 µm wide, but sometimes weakly developed. Remainder of duct 150–280 µm long. Spermathecal duct narrows near ectal end, but is not convoluted as in typical specimens ( Figs. 7 View FIGURE 7 A–C, 10C). Spermathecal ampulla 200–800 (350) µm long; may be as described for typical E. pacificus , but usually less elongate. In most specimens the epithelial cells are not thickened or vacuolated entally, and the sperm cells are usually distributed throughout the ampulla in a loose bundle ( Fig. 10 View FIGURE 10 D).
Variant 2
Length of preserved worms 19–25 mm, 78–98 segments; width 0.40–0.65 mm in X. Blind lateral blood vessels well developed in posterior segments: from about XXX to about L there are anterior and posterior groups of 1–3 short, digitiform vessels; posterior to L, two pairs of lateral vessels have 4–6 (or more) long branches ( Fig. 2 View FIGURE 2 E).
Male pore on a short papilla, as described for Variant 1 specimens; epidermal pad surrounding pore 86–145 µm wide ( Fig. 8 View FIGURE 8 C). Atrial duct 360–1030 (690) µm long; ampulla length 360–740 (560) µm, maximum diameter 89–132 µm. Poor specimen fixation limits histological observations, but longitudinal muscle layer of atrium appears to be 10–19 µm thick, inner circular muscle 4–7 µm. Longitudinal muscle appears to be parallel to longitudinal axis of ampulla. Maximum diameter of atrial lumen 0.41–0.66 (0.55) times ampulla diameter. Anterior male funnels smaller than posterior, usually with sperm.
Spermathecal morphology similar to Variant 1, but placement of spermathecal pores varies. In the material collected in 2002, five specimens have paired spermathecal pores anterior to ventral chaetae, usually in the secondary annulus groove in IX; an additional pore is near the midventral line, posterior to the chaetae ( Fig. 3 View FIGURE 3 B, 7D). Six specimens have a single, midventral spermathecal pore, posterior to or between the ventral chaetal bundles; one worm has a single anterolateral pore in addition to the posteromedian one. Anterior pores inconspicuous, but posteromedian pores usually more prominent, on low mounds. Ectal vestibule of spermathecal duct 70–120 µm long; remainder of duct 140– 240 µm long. Spermathecal ampulla nearly spherical to elongate, 210–730 µm long; histology variable. Posteromedian spermathecal ampulla often larger than anterior ones (if present), extending into X or XI ( Fig. 7 View FIGURE 7 E–F). Sperm usually present in all ampullae.
Specimens from the Royal Ontario Museum are highly contorted, in poor condition. Four appear to have paired, anteriorly placed pores; three also have midventral pores posterior to the chaetal bundles; and one has a single midventral pore. The single specimen from a spring near the Nestucca River has the 3spermatheca arrangement ( Fig. 7 View FIGURE 7 D).
Remarks
The elongate, petiolate atria with numerous small prostate glands, and the very thin vasa deferentia, embedded in the thick, muscular wall of the atrium, resemble those of other Eclipidrilus species. As is usual for the genus, E. pacificus has reduced (although functional) male funnels associated with small anterior testes in IX. Morphology of the pharynx, blood vessels and nephridia are also typical for the genus (see discussion).
Median, unpaired male pores also occur in Eclipidrilus asymmetricus ( Smith, 1896) and E. lacustris , but neither of those species has laterally paired spermathecal pores on the chaetal lines, anterior to the chaetae. Although the general proportions of the atrium and the accessory glands at the male pore are similar to those of E. pacificus , E. asymmetricus is described as having a conspicuous proboscis and median, longitudinally paired spermathecal pores. Eclipidrilus asymmetricus is also described as lacking anterior vasa deferentia, male funnels on 9/10, and testes in IX, but most other characters related to the reproductive organs are inadequately described for comparison. Unfortunately, the single mature specimen used in the original description has disappeared, and a subsequent search of the type locality was unsuccessful ( Wassell 1984).
Typical E. pacificus specimens are distinguished from all variant populations by a penis formed of extruded atrial duct lining, and by the larger spermathecal vestibules. The atrium, the concave, epidermal pad and the accessory glands surrounding the male pore also are much larger and well developed, and the musculature surrounding both spermathecal and atrial ducts is also more extensive. The penis appears structurally similar to that of E. frigidus , but it is much more extensively developed in the latter species (see below). In the variant populations, the penis is reduced to a small papilla without obviously extrudable lining cells. The occurrence of a posteromedian spermathecal pore in IX usually distinguishes the Variant 2 specimens, from the lower Columbia River and one other site, but the variable number (1–3) of spermathecae makes it difficult to define limits that would indicate formal taxonomic status. A similar problem was noted for Eclipidrilus ithys Brinkhurst, 1998 , in which atria may be paired or single. Eclipidrilus pacificus specimens from the southernmost Washington collection site (South Naselle River) may be intermediate between the typical form and Variant 1: they resemble the typical Olympic Peninsula material except that the lining cells of the penial structure are not as highly developed and elongate.
The variants may correspond to geographic regions. Typical E. pacificus have been collected only in coastal drainages of Washington, with most records from the Olympic Peninsula. Variant 1 is relatively widespread, with several records from the Coast Ranges of Oregon, two from small tributaries to Puget Sound in northern Washington, and two from northern Idaho. The wide but disjunct distribution of Variant 1 records precludes morphometric analysis that could rule out clinal variation in characters. An additional problem is that specimens from some sites were not well fixed, leading to possible artifacts. Given the persistent ambiguity in species limits between E. palustris and Eclipidrilus daneus Cook, 1966 (see below), it seems best at this point to consider all E. pacificus populations as representatives of a single, variable species.






| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Eclipidrilus pacificus
| Steven & Fend, V. 2005 |
Eclipidrilus ithys
| Brinkhurst 1998 |
Eclipidrilus daneus
| Cook 1966 |