
Drosera margaritacea T.Krueger
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.501.1.2 |
|
persistent identifier |
https://treatment.plazi.org/id/038787B0-876A-FF94-B4DF-C716FE43F928 |
|
treatment provided by |
Marcus |
|
scientific name |
Drosera margaritacea T.Krueger |
| status |
sp. nov. |
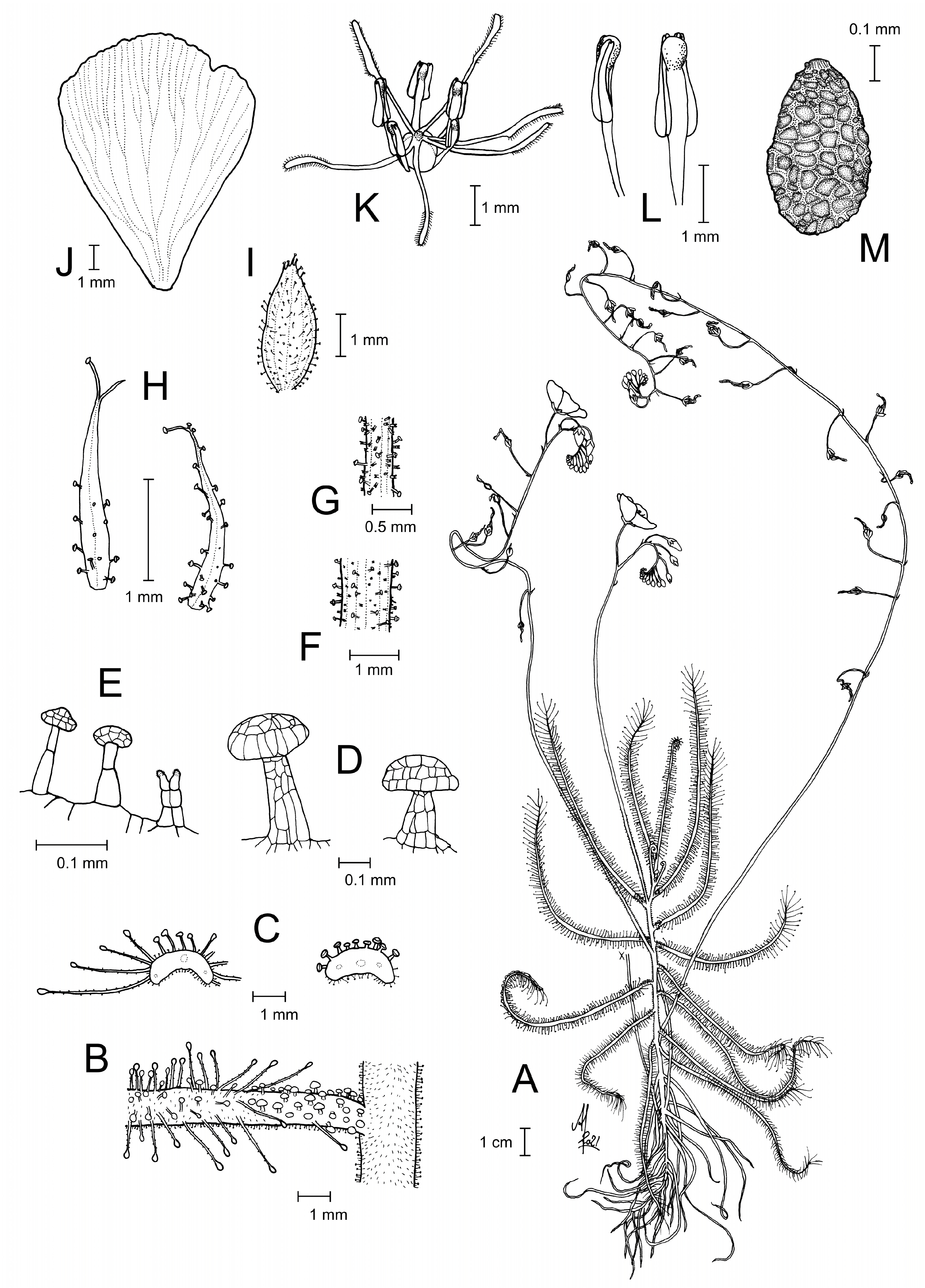
1. Drosera margaritacea T.Krueger View in CoL & A.Fleischm., sp. nov. ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 + 6–9)
Type: — AUSTRALIA. Western Australia: Kimberley. 60 m N of Great Northern Hwy., 80.4 km ESE of intersection with Derby Hwy., 123 km WNW of Fitzroy Crossing, narrow drainage channel, reddish-brown sandy clay, 18 July 2020, T . Krueger 7 (holotype PERTH!, isotype MEL!) .
Diagnosis:— Drosera margaritacea is morphologically similar to Drosera finlaysoniana Wall. ex Arn. in Hook. from which it differs (contrasting characters in parentheses) by its petiolate leaves, borne on a 1.5–3.5 mm long petiole (leaves sessile), petiole covered with 20–50 stalked secretive glands 0.10–0.40 mm long with a multicellular stalk and translucent milky white, hemispherical to depressed ovoid gland head 0.10–0.30 mm in diameter that remains present even in dried material (petiole absent), stem, leaves, peduncle, pedicels and sepals covered with 0.05–0.30 mm long stalked capitate trichomes with uniseriate stalk and yellow, spherical, non-secretory gland head ca. 0.05 mm in diameter (stem, abaxial leaf surface, peduncle, pedicels and sepals covered with 0.1–0.3 mm long secretory capitate trichomes with uniseriate stalk and flattened gland head secreting a translucent mucilage droplet 0.1–0.2 mm in diameter), T-or Y-shaped trichomes absent (adaxial leaf bases and margins covered with 0.05–0.40 mm long T-or Y-shaped trichomes), inflorescence (20–)30–55(–65) cm long, with at least 30–50 flowers, its peduncle held upright, ± parallel to stem and much exceeding the carnivorous leaves (inflorescence (3–)6–18(–26) cm long, (2–)6–20(–26)- flowered, peduncle usually horizontal [this character is typically very obvious in live specimens, but sometimes not easily discerned in pressed herbarium material], shorter than or at most equalling the leaves in length), flowers 18–24 mm in diameter (7–14(–20) mm in diameter), stamen connective longer than filament, shorter than thecae and with rounded yellow tip ca. 0.2–0.4 mm in diameter (stamen connective usually much shorter than filament, as long as the thecae, lacking a rounded yellow tip).
Description:— Annual herb, 25–50(–90) cm tall including inflorescence. Roots simple, short, slightly succulent, terete, poorly branched. Stem upright or ascending, 8–16(–20) cm long, simple (unbranched), terete, 1.2–2.5 mm in diameter, green, turning reddish with age, internodes (1.5–)2.0–3.0(–8.0) mm long in flowering specimens; stem with indumentum consisting of two different types of trichomes: 0.05–0.15 mm long translucent, biseriate (double-tipped) trichomes (= type 5 sensu Länger et al. 1995) and 0.05–0.30 mm long stalked capitate trichomes with uniseriate stalk and yellow, spherical, non-secretory gland head ca. 0.05 mm in diameter. Leaves filiform, (40–)50–100(–130) mm long in flowering-sized specimens, straight, vernation circinate, sometimes emitting a strong honey-like scent; freshly developed leaves held erect or patent and with slightly curved lamina, leaves held horizontal and lamina straight with age, senescent leaves persistent, patent or reflexed towards the stem; stipules absent; petiole linear, 1.5–3.5 mm long, 1.5–2.4(–3.0) mm wide at its greatest width (at transition to the lamina), green, in section depressed narrowly reniform with slightly reflexed lateral margins; petiole adaxial surface and margins with 20–50 distinctive, stalked secretive glands with multicellular stalk (0.1–0.4 mm long) and translucent milky white, hemispherical to depressed ovoid gland head, 0.1–0.3 mm in diameter, turgescent and adhesive in living specimens, maintaining its shape but losing stickiness in dried material; petiole adaxial surface additionally very sparsely covered with 0.05–0.15 mm long, translucent, biseriate, double-tipped trichomes like those present on the stem (but yellow-headed capitate trichomes absent on adaxial petiole surface); abaxial petiole surface indumentum identical to the stem indumentum; lamina narrowly linear-lanceolate with long acuminate tip, (38–)48–100(–120) mm long (excluding tentacles), 1.5–2.4(–3.0) mm wide (excluding tentacles) at its greatest width; lamina yellowish-green, its adaxial surface covered with stalked, carnivorous, secretive capitate glands (tentacles) 1–5 mm long, tentacle stalk translucent white or tinged reddishpink, with regular, hemispherical microscopic projections, tentacle gland head red or orange-yellow, all tentacles with radially symmetric gland head; additionally, lamina adaxial surface and margins sparsely covered with stalked milkywhite glands of the same type as present on the petiole adaxial surface (slightly smaller on the lamina: gland stalk 0.10– 0.30 mm, hemispherical gland head 0.15–0.20 mm in diameter) and with 0.05–0.15 mm long, biseriate, double-tipped trichomes and 0.05–0.30 mm long yellow-headed capitate trichomes; indumentum of lamina abaxial surface identical to stem indumentum. Inflorescences (1)3–6, each forming a many-flowered (at least 30–50 flowers per peduncle), bracteate scorpioid cyme; inflorescence indumentum (covering peduncle, rhachis, bracts, pedicels, calyx and sepals) identical to stem indumentum; young inflorescences yellowish-green or bronze, turning reddish with age; scape (200–)300–550(–650) mm tall including peduncle; peduncle (80–)90–160(–190) mm long, much exceeding the leaves in length, terete, 1.1–1.5 mm in diameter, ascending vertically (± parallel to the stem axis in young inflorescences, mostly held at an angle <45° from the stem in older inflorescences and at fruit, rarely diverging up to 90° from stem); fruiting pedicels spaced by 5–18(–30) mm; pedicels terete, 0.2–0.6 mm in diameter, 9–25(–35) mm long in fruit; pedicels straight upright at anthesis becoming more or less arcuated or reflexed in fruit (arcuated horizontal/slightly downwards from the inflorescence axis, then sharply curving upright at the apex, so that the seed capsule is held upright). Bracts subulate, 1.0–2.5(–3.0) × 0.05–0.20 mm, persistent, margins entire, apex acute, shallowly truncate or bifid, often ending in a glandular trichome, adaxial surface glabrous, abaxial surface indumentum identical to the stem indumentum. Flowers 18–24 mm in diameter. Sepals 5, basally adnate (usually not overlapping in developing buds), lanceolate-elliptical, margins serrate or entire, 2.0– 3.5 mm long, 0.9–1.4 mm at their greatest width, abaxial surface with indumentum identical to the stem indumentum. Petals 5, obovate to broadly obovate, 9–12 mm long, 7–9 mm at their widest point, apical margin slightly irregularly crenulate to undulate, petals overlapping near their base, petals bright pink. Stamens 5, narrowly lanceolate, 2.5–3.0 mm × 0.7–0.9 mm; filaments linear, 0.9–1.5 × 0.2–0.5 mm, white; anthers bithecate, exceeding (or at least equalling) the filament in length, 1.4–2.0 mm long, connective dilated, narrowly triangular, 0.5–0.6 mm wide, slightly shorter than the thecae, with rounded, swollen yellow apex ca. 0.2–0.4 mm in diameter; thecae pale yellowish-white; pollen pale yellowish-white. Ovary 3-carpellate, fused, subglobose, 1.0– 1.5 mm in diameter, glabrous, pale green. Styles 3, each basally divided into 2 entire style-arms; style-arms translucent white, 1.5–3.0 mm long (excluding stigmatic portion), flagelliform, terete, ca. 0.1–0.2 mm in diameter, glabrous, positioned horizontally, graduating into stigmatic part; stigmas 1.2–2.2 mm long, translucent white, apex slightly (rarely strongly) curved upwards, stigmatic surface papillate. Seeds numerous, (350–)400–500 × (200–)250–300 µm, ellipsoid to broadly ellipsoid or ovoid, testa black or grey, testa surface more or less isodiametrically reticulate, with anticlines comparatively thick and raised, periclinal walls tabular, covered with microscopic, bladed wax crystals.
Etymology: —The species’ epithet (from Latin margaritaceus = pearly, pearl-bearing) refers to the indumentum of unique, stalked secretive glands with translucent milky white, hemispherical to depressed ovoid (pearl-like) gland heads, as well as to the characteristic swollen, roundish (pearl-like) apical dilatation of the anther connective.
Taxonomic notes: — Drosera margaritacea is morphologically most similar to D. finlaysoniana as both species share a similar habit (at least of the vegetative parts) with leaves that appear glandular to the very base (no petioles apparent), similar seed shape and scent production. However, in D. finlaysoniana [as well as in D. aquatica Lowrie (2014: 1268) which is frequently confused with D. finlaysoniana , see Discussion: Taxonomy] it is the carnivorous tentacles that reach the leaf base, hence that species is lacking a petiole, while in D. margaritacea , the petiole is short but clearly apparent upon close examination and covered by distinctive glands with multicellular stalks and translucent to milky-white, depressed heads. This gland type /trichome type is not paralleled in any other member of D. section Arachnopus , nor any other Drosera species. Drosera aquatica is additionally separated from D. margaritacea and D. finlaysoniana by its stem and inflorescence indumentum which consists of long, patent, simple, translucent eglandular hairs ( Lowrie 2014).
The comparatively long and many-flowered, vertically arising inflorescence readily allows identification of Drosera margaritacea in situ as well as in herbarium material, and even in low-resolution scans of herbarium specimens. A similar inflorescence habit is found only in D. barrettiorum Lowrie (2014: 1269) and in large “variants” of D. hartmeyerorum and D. serpens from the Kimberley (T. Krueger pers. obs.). Indeed, D. hartmeyerorum is the only other species in D. section Arachnopus whose anthers exhibit similar, swollen, apical connective dilatations. However, in D. hartmeyerorum this apical connective swelling of the anthers is white (rarely yellow) and much exceeding the thecae ( Barrett & Lowrie 2013, Lowrie 2014, T. Krueger & A. Fleischmann pers. obs.), while in D. margaritacea the yellow, swollen part of the connective is subequal to the thecae in length. In the anthers of D. barrettiorum , the deltoid connective is much dilated and curved to the ventral side, resulting in a yellow, dorsal swelling, the anthers of this species are subequal to (slightly exceeding) the connective. While D. serpens is readily distinguished from D. margaritacea by its well-expressed petioles, D. barrettiorum and D. hartmeyerorum always produce groups of relatively large and conspicuous yellow emergences on their adaxial petiole surfaces, and thus can readily be told apart from all remaining members of D. section Arachnopus ( Lowrie 2014) .
Distribution and habitat: — Drosera margaritacea is endemic to the western part of the Kimberley region of tropical northern Western Australia, apparently limited to lowlands of the Canning Basin, with records made at elevations of 10–70(–180) m above sea level ( Fig. 3A View FIGURE 3 ). It is known from the base of the Dampier Peninsula in the west to Christmas Creek Homestead (ca. 95 km SE of Fitzroy Crossing) ca. 400 km to the east, extending northwards to Derby and throughout the Yampi Peninsula ( Fig. 3A View FIGURE 3 ). Although not reported from there yet, it is likely that D. margaritacea also occurs in central or northern parts of the Dampier Peninsula, as this area is in close geographic proximity to the Yampi Peninsula and features suitable habitat as well as similar climate and geomorphology. A single PERTH record with geocoordinates given for near Kununurra in the eastern Kimberley (A. Lowrie 2461) has been determined to originate from the Yampi Peninsula, as it was evidently collected by Allen Lowrie during his 2000 expedition to that area ( Lowrie 2001).
The morphologically similar (and putatively closely related) D. finlaysoniana , while abundantly recorded in southern, eastern, and central parts of the Kimberley, is not known to occur within the distribution range of D. margaritacea , indicating that the two species may be allopatric ( Fig. 3A View FIGURE 3 ). In the Christmas Creek area, however, both species apparently occur within a distance of less than 40 km from each other. Additionally, D. finlaysoniana is apparently limited to higher elevations (above ca. 200 m) in the central and western parts of the Kimberley, i.e. the area of parapatry of the two species ( Fig. 3A View FIGURE 3 ).
Drosera margaritacea grows in sandy soils near the edges of seasonally wet swamps, creeks, and seepage areas, usually amongst dense grassy vegetation. Associated flora includes Fuirena ciliaris (L.) Roxb. ( Cyperaceae ; pers. obs. and mentioned on specimen label of R. Jensen & J.E. Kemp RJ 3983), Cyperus spp. ( Cyperaceae ; pers. obs. and also mentioned on L. Wallis LW 97A/112), Schoenoplectiella dissachantha (S.T.Blake) Lye ( Cyperaceae ; mentioned as “ Schoenoplectus dissachanthus ” on L. Wallis LW 97A/112), Eriachne festucacea F.Muell. ( Poaceae ; mentioned on R. Jensen & J.E. Kemp RJ 3983), Calandrinia sp. ( Montiaceae ; pers. obs.) and Melaleuca sp. ( Myrtaceae ; pers. obs. and mentioned on L. Wallis LW 97A/112).
Ecology and Phenology: —Annual. Flowering has been recorded from May to August, i.e. during the northern Australian dry season. However, the phenology of some examined herbarium specimens collected in early May (K. Coate 406, A. Lowrie 2216, A. Lowrie 2219), and of individuals studied in situ in mid-August (T. Krueger pers. obs. 2014), indicates that flowering can occur from April to at least early September. Following particularly dry wet seasons, this species has been observed to be completely absent from known locations even at anthesis time (T. Krueger pers. obs. 2020). Therefore, it is possible that seed germination in this species strongly depends on the amount of wet-season flooding.
Conservation status: —Least Concern (LC) according to IUCN Red List Criteria ( IUCN 2012). Drosera margaritacea is known from at least 15 locations as defined by IUCN Red List guidelines ( IUCN 2012), covering a wide area of the Kimberley. None of these sites are situated on conservation managed lands (CALM Act lands). While commonly localised and occurring in rather small populations (two populations, one with 5 and one with ca. 100 individuals, were observed by the first author in 2012, 2014 and 2020), very large populations are known to exist at some sites (e.g., at Lanlacatta Swamp, where it is the “most abundant insectivorous plant in the area” according to specimen label data of L. Wallis LW 97A/131). At present, the species does not meet any criteria for conservation code listing in Western Australia, but further surveys to assess its wider distribution and potential threats (such as cattle farming and altered fire regimes) are recommended.
Additional specimens examined (paratypes):— AUSTRALIA. Western Australia: [Kimberley region], Christmas Creek , May 1927, A. J . Ewart s.n. ( PERTH 3289516 About PERTH !, PERTH 3289095 About PERTH !); W. A . Kimberleys [sic.!], 3 km E of Stuart River, Oobagooma Road , between Stuart and Robinson Rivers , c. 74 km NNE of Derby, 14 June 1976, A. C . Beauglehole 52915 (DNA D0269214 photo!); Prior’s Bore, ca 10 km N of Great Northern Highway , at a point 135 km E of Broome along Broome-Derby Road , damp soil, 04 July 1982, K. F . Kenneally 8538 ( PERTH 3289451 About PERTH !, CANB); Deep Creek, 55 km E of Broome on Great Northern Highway, Kimberley , in damp sand, 29 June 1993, K. F . Kenneally 11379 ( PERTH 3048292 About PERTH !, CANB); W Kimberley coast, Jinunga River off The Graveyard , seepage areas beside pool, 02 July 1996, K. F . Kenneally 11667 ( PERTH 4410521 About PERTH !); W Kimberley coast, Kyulgam River, a freshwater creek off Strickland Bay , on creek edge in damp soil, 03 July 1996, K. F . Kenneally 11686 ( PERTH 4410742 About PERTH !); Dugong Falls, Talbot Bay , top of falls, 08 July 1996, K. F . Kenneally 11758 ( PERTH 4411404 About PERTH !); Near Blina on Great Northern Highway, Kimberley , 13 May 1997, K . Coate 406 ( PERTH 4942795 About PERTH !, MEL); c. 1 km from Taylors Lagoon, Broome, growing in sandy soil, 26 June 1997, A . Lowrie s.n. ( PERTH 6440355 About PERTH !); Blina Swamp on Blina Station, edge of Blina Swamp , 23 July 1997, L . Wallis LW 97 A/112 ( PERTH 5025540 About PERTH !); Lanlacatta Swamp on Camballin Station, Kimberley , Edge of Lanlacatta Swamp , 27 July 1997, L . Wallis LW 97 A/131 ( PERTH 5025745 About PERTH !); c. 129 km W of Fitzroy Crossing, Kimberley , 09 May 1999, A . Lowrie 2216 ( PERTH 6440576 About PERTH !); c. 129 km W of Fitzroy Crossing, Kimberley , 09 May 1999, A . Lowrie 2219 ( PERTH 6440568 About PERTH !); Crossing Falls [Yampi Peninsula], Kimberley , 30 July 2000, A . Lowrie 2461 ( PERTH 6440274 About PERTH !); Sand pit, 11.2 km E on the Gibb River Road from the Broome / Derby junction outside Derby townsite, flat, red sand, freshwater ponds, 12 June 2006, L. S. J . Sweedman 6853 ( PERTH 7640277 About PERTH !); Stewart River where Kimbolton Road crosses, c. 9.4 km ESE of Kimbolton homestead, Yampi Training Area , on sand and cobble in river channel, 14 July 2018, R . Jensen & J. E . Kemp RJ 3983 ( PERTH 9201726 About PERTH !) .
Georeferenced photographs: — AUSTRALIA. Western Australia: Fitzroy Crossing Road, Kimberleys [sic.!] , Western Australia, 23 June 2017, R. Fryer & J. Newland 6177 (northqueenslandplants.com); Taylor’s Lagoon, Willare , Western Australia, 24 June 2017, R. Fryer & J. Newland 6190 (northqueenslandplants.com) .
Additional localities reported by Lowrie (2001): — AUSTRALIA. Western Australia: 8 km NE of Karriwell Yard [Yampi Peninsula], open grassy plain, July 2000 ; 11 km NE of Karriwell Yard [Yampi Peninsula], near minor seepage area, July 2000 .


| N |
Nanjing University |
| T |
Tavera, Department of Geology and Geophysics |
| PERTH |
Western Australian Herbarium |
| MEL |
Museo Entomologico de Leon |
| A |
Harvard University - Arnold Arboretum |
| J |
University of the Witwatersrand |
| W |
Naturhistorisches Museum Wien |
| E |
Royal Botanic Garden Edinburgh |
| C |
University of Copenhagen |
| K |
Royal Botanic Gardens |
| F |
Field Museum of Natural History, Botany Department |
| CANB |
Australian National Botanic Gardens |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| S |
Department of Botany, Swedish Museum of Natural History |
| R |
Departamento de Geologia, Universidad de Chile |
| NE |
University of New England |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.