
Discothyrea patrizii Weber, 1949
|
publication ID |
https://doi.org/10.1093/isd/ixz015 |
|
DOI |
https://doi.org/10.5281/zenodo.5922626 |
|
persistent identifier |
https://treatment.plazi.org/id/03D9AC4A-E56F-FF86-FF64-F9A1BA0F04DC |
|
treatment provided by |
Plazi |
|
scientific name |
Discothyrea patrizii Weber, 1949 |
| status |
|
Discothyrea patrizii Weber, 1949 View in CoL
( Figs. 4N View Fig , 6N View Fig , 7N View Fig , 8N View Fig , 9N View Fig , 10N View Fig , 11N View Fig , 12N View Fig , 14N View Fig , 45,
46 View Fig , 47 View Fig ; Supp Video S14 [online only])
Discothyrea patrizzii Weber, 1949: 2 . [Justified emendation of spelling to Discothyrea patrizii View in CoL by Brown, 1958a].
Type Material
HOLOTYPE, pinned queen, KENYA, high East African plains , densely wooded donga or ravine, floor cover of leaves and humus, 1670 to 1770 m, 5.II.1948 [ holotype presumably lost, not at MCZC] . PARATYPE, pinned worker, KENYA, Nairobi , [ −1.317, 36.763], ca. 1750 m, 1945 ( Patrizi) ( MCZC: MCZ_Type_32184) [examined] GoogleMaps .
Virtual dataset. Volumetric raw data (in DICOM format), 3D rotation video, still images of surface volume rendering, and 3D surface (in PLY format) of a nontype specimen (CASENT0235472) in addition to stacked digital color images illustrating head in full-face view, profile and dorsal views of the body.The data are deposited at Dryad (Hita Garcia et al. 2019, http://doi.org/10.5061/dryad.3qm4183) and can be freely accessed as virtual representation of the species. In addition to the data at Dryad, we also provide a freely accessible 3D surface model at Sketchfab (Model 14).
Nontype Material
KENYA: Nairobi , −1.317, 36.763, ca. 1750 m, soil sample, 4.XI.1974 ( V. Mahnert) GoogleMaps ; TANZANIA: Kilimanjaro Region, Kindoroko Forest Reserve , −3.74520, 37.64267, 1739 m, primary forest, leaf litter, Winkler, 5.–8.IX.2005 ( P. Hawkes, J. Makwati, R. Mtana) GoogleMaps ; Kilimanjaro, Mt . Kilimanjaro, −3.16699, 37.23584, 1920 m, montane forest, 20.X.–10.XI.2011 ( M. Peters) GoogleMaps ; Kilimanjaro, Mt. Kilimajaro , −3.186, 37.2546, 1640 m, farmland near montane forest, 1.XI.2011 ( A. Mayr) GoogleMaps ; Kilimanjaro, Mt . Kilimanjaro, −3.2604, 37.4180, 1620 m, montane forest, 12.XII.2011 ( M. Peters) GoogleMaps ; Iringa Region, Ndudulu Forest Reserve, −7.78912, 36.48539, 1567 m, primary forest, hand collected, 23.–26.X.2007 ( P. Hawkes, M. Bhoke, U. Richard); Mkomazi Game Reserve, forest above Ibaya, −3.96667, 37.78333, montane forest, Winkler bag extraction from sifted leaf litter from fern-covered rock, 27.XI.1995 ( H.G. Robertson); Mkomazi Game Reserve , Kinondo forest , −3.91667, 37.76667, Model 14. 3D surface model of D. patrizii Weber, 1949 (CASENT0235472). An interactive version of this model is available in the HTML version of this article online and at https://sketchfab.com/3d-models/9fe9753fa49a435db58 38fb0d6486dbf GoogleMaps .
montane forest, Winkler bag extraction ex leaf litter, 9.V.1996 ( H.G. Robertson); Mkomazi Game Reserve, Maji Kununua, −3.88333, 37.81667, 1600 m, montane forest, Winkler bag leaf litter extraction, 7.XII.1995 ( H.G. Robertson); Morogoro Region, Mamiwa- Kisara Forest Reserve, −6.37530, 36.93711, 1989 m, primary forest, leaf litter, Winkler, 16.–21.VIII.2005 ( P. Hawkes, J. Makwati, R. Mtana); Morogoro Region, Mamiwa-Kisara Forest Reserve, −6.37530, 36.93711, 1989 m, primary forest, pitfall trap, 16.–21. VIII.2005 ( P. Hawkes, J. Makwati, R. Mtana); South Pare Forest, −4.130556, 37.88389, montane forest, Winkler bag leaf litter extraction, 29.XI.1995 ( H.G. Robertson); South Pare Mountains, −4.13333, 37.88333, 1598 m, montane forest, Winkler bag extraction ex leaf litter, 18.IV.1996 ( S. van Noort ); Tanga Region, Nilo Forest Reserve, −4.91456, 38.67712, 1006 m, primary forest, hand collected, 1.–4.IX.2005 ( P. Hawkes, J. Makwati, R. Mtana); West Usambara Mountains, Shume Gologolo, −4.70, 38.23, disturbed montane forest, Winkler bag extraction ex leaf litter, 13.V.1996 ( H.G. Robertson); West Usambara Mountains, Site 2, −4.733, 38.25, 2001 m, montane forest, Winkler bag extraction ex leaf litter, 1.V.1996 ( M. Stander, S. van Noort ).
Diagnosis
The following character combination distinguishes D. patrizii from the remainder of the complex: standing pilosity absent from mesosoma and abdominal terga; propodeum without strong angles or denticles; eyes present, relatively large, and round (OI 5–8); in dorsal view mesosoma relatively slender (DMI 52–58; DMI2 78–87) and moderately narrowed posteriorly; in profile mesosomal outline comparatively flat; mesotibia without apicoventral spur; petiolar node strongly attenuated dorsally (DPeI 314–489; LPeI 314–467); sculpture generally reduced, declivitous face of propodeum without foveolae; color highly variable: unicolorous matte orange to very deep brown with yellowish appendages.
Worker Measurements and Indices ( n = 15)
EL 0.03–0.04; HL 0.38–0.54; HW 0.30–0.47; SL 0.19–0.30; PH 0.19–0.28; DML 0.26–0.39; PW 0.23–0.33; PrH 0.23–0.34; WL 0.40–0.57; HFL 0.22–0.35; PeL 0.04–0.07; PeW 0.16–0.22; PeH 0.16–0.22; LT3 0.28–0.40; LT4 0.24–0.38; OI 5–8; CI 78–87; SI 50–58; LMI 44–50; DMI 52–58; DMI2 78–87; ASI 85–98; HFI 54–63; DPeI 314–489; LPeI 314–467.
Worker Description
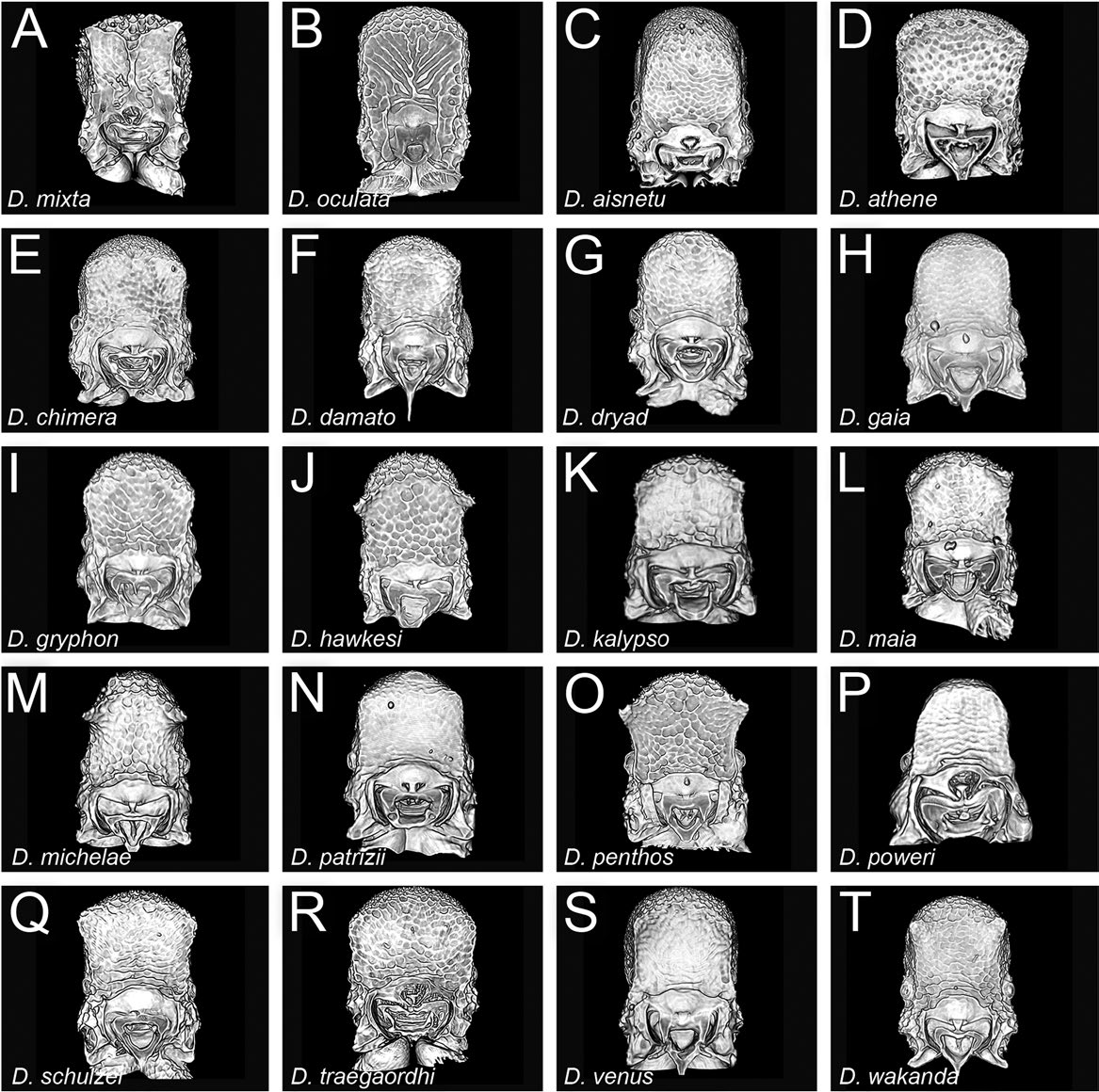
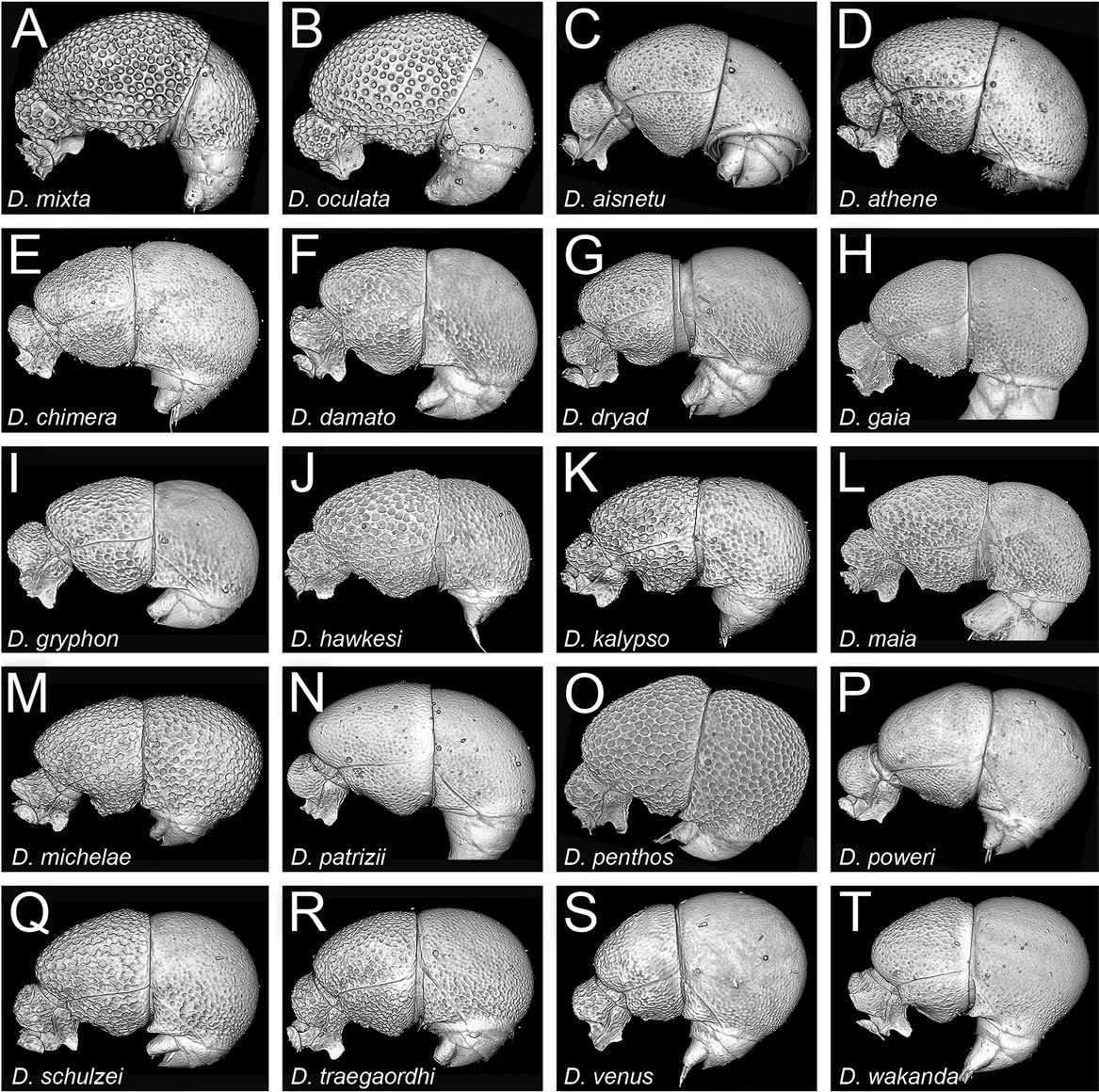
Head conspicuously longer than broad (CI 78–87), posterior head margin straight to convex, posterodorsal corners of head broadly rounded; sides of head in frontal view gently subparallel to convex posterad eyes, very slightly concave between eyes and anterolateral corner of gena; eyes present, relatively large (OI 5–8), round, comprising several ommatidia, placed about a third of the way between anterolateral corner of gena and posterior head margin; eyes visible in frontal view; frontal lamella fairly short and triangular in profile, apex rounded to acute; lamella more or less evenly translucent across its disc, sometimes with a thinner basal spot, but lacking a distinct fenestra; medial clypeus convex, lateral clypeus curving broadly between antennal sockets and anterolateral corners of head, bearing short, curved setae. Antenna with shorter to moderately long scape (SI 50–58), scape slightly expanded apically, very gently bent; pedicel campaniform, slightly longer than broad; true antennomere count eight; apparent antennomere count seven to ten, flagellomeres basad apical club highly compressed, taken together only about as long as apical club. Ventral head with narrowly carinulate, V-shaped preoccipital ridge without anteromedian carina; medial area of hypostoma triangular, arms slightly narrowed, similar in width across their length; palpal formula not examined. Mandible edentate except for small, curved prebasal denticle; occasionally a slight preapical swelling present; basal angle rounded; ectal face with weak carina running from about midway between basal angle and prebasal denticle or mandibular apex, becoming confluent with masticatory margin around halfway along its length, leaving a short, comma-shaped depressed area.
Mesosoma sloping posteroventrally, pronotum clearly higher than propodeum; in dorsal view mesosoma relatively slender (DMI 52–58; DMI2 78–87) and moderately narrowed posteriorly, pronotum somewhat wider than propodeum; pronotal humeri rounded; posterior propodeal margin straight; posterodorsal corners of propodeum rounded angulate but not denticulate; declivitous face of propodeum slightly to moderately concave in profile and oblique posterior view; propodeal spiracle small but sometimes distinct (more so in darker morphs), directed posterolaterally; propodeal lobes well-developed, flangelike.
Legs short to intermediate in length (HFI 54–63) and somewhat narrow; mesotibia without apicoventral spur; mesobasitarsus fairly short, subequal in length to tarsomeres II–IV taken together.
Petiolar node attenuated dorsally, somewhat variable in shape, about 2.9 to 3.8 times higher than long (LPeI 286–383); in profile anterior face of node convex, apex peaked, somewhat rounded to subacute, posterior face posteroventrally sloping to subvertical; in dorsal view, node rectangular, about 3.0 to 3.8 times broader than long (DPeI 300–383), sides subparallel to slightly divergent posteriorly; in anterior view, petiolar outline pentagonal, angles well-rounded; in oblique anterodorsal view, anterior face flat; subpetiolar process variable in shape, dentate to broadly lobate, apex acute to rounded.
Abdominal segment 3 campaniform, widest just anterad end of segment; tergite slightly anteriorly prolonged over petiole; sternite evenly curved to posteriorly bulging in profile (deepest point at around or slightly posterad longitudinal midline); AS 3 without median ridge, without carinate prora, but anterior face still depressed, anterior margin of ventral face flat to weakly concave in ventral view; AT3 and AT4 approximately equal in length, or AT4 only slightly (about 1.2 times) longer than AT3 (ASI 100–115); AT4 gently recurved, hemidemispherical; AS 4 with poorly developed anterior lip, overlapping slightly more than half the width of AS 3, anterior margin straight in ventral view; successive abdominal segments short, telescopic, often concealed.
Sculpture on head, mesosoma, and petiole very finely, shallowly punctulate, often appearing colliculate or even scabriculous in darker morphs; punctulae on lateral mesosoma becoming reticulate to substrigulate; declivitous face of propodeum predominantly smooth; mandibles similarly sculptured to head, somewhat shining between piligerous punctulae; abdominal segment 3 with coarser, more distinct punctae, similarly distributed on tergite and sternite; AT4 with minute but distinct, very densely arranged piligerous punctae, clearly shinier than AT3.
Setation mostly consisting of appressed white pubescence, of similar density on head and mesosomal dorsum, generally longer and denser on gastral terga, more dilute on lateral mesosoma and lateral portions of abdominal terga; setae on head often inclined towards the longitudinal midline; ectal face of mandible with moderately long, fine, appressed to decumbent setae; masticatory margin with row of straight, stout setae; occasionally a few short decumbent to erect hairs present on dorsal surfaces, particularly of AT3; appearance of setation highly variable between color morphs: in light individuals pubescence inconspicuous, while very distinct on dark individuals; abdominal segments 5 through 7 with long, flexuous standing setae. Appendages with well-developed, evenly distributed appressed pubescence.
Color highly variable: unicolorous matte orange to very deep brown with yellowish appendages.
Distribution and Biology
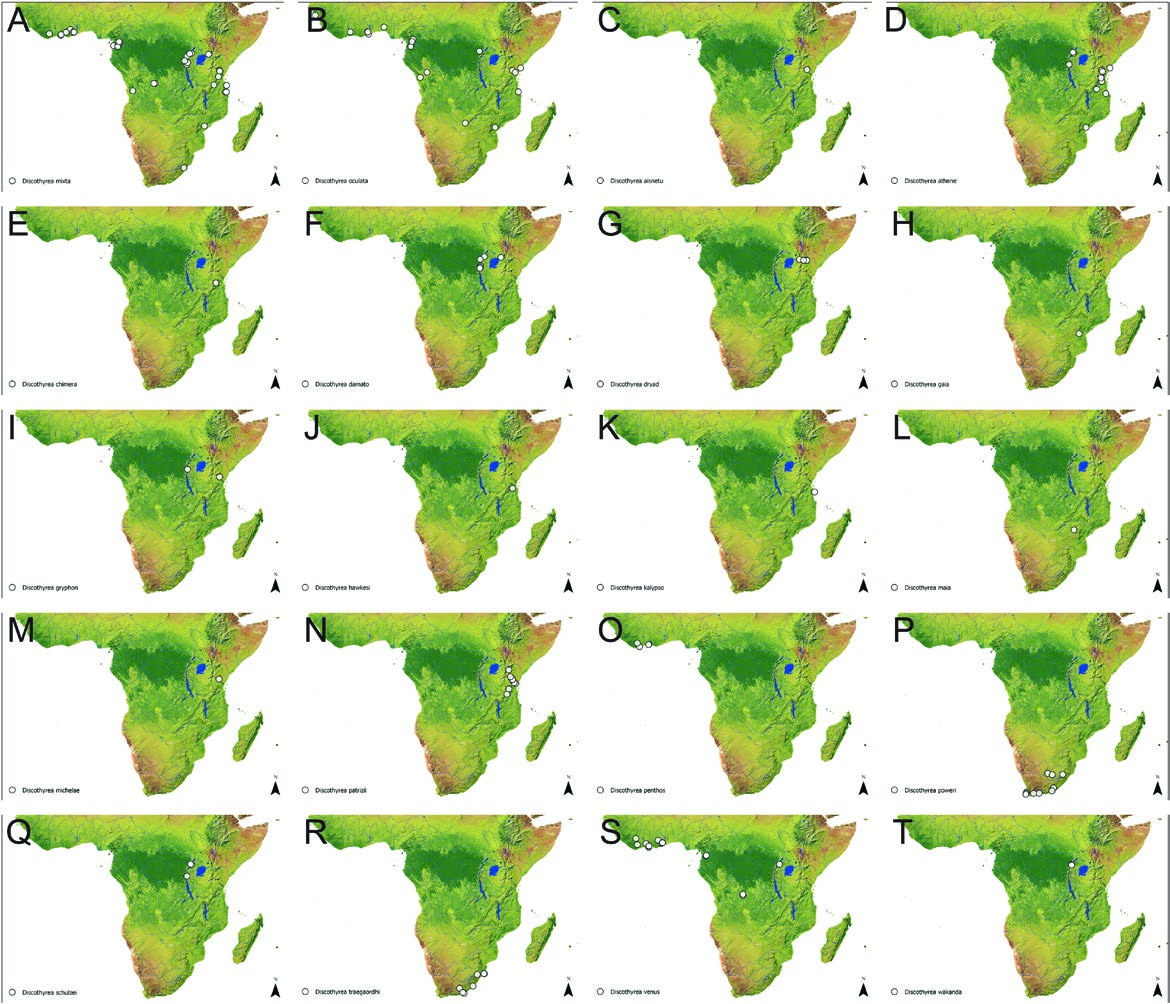
Discothyrea patrizii is known from montane forests in Kenya and Tanzania ( Fig. 4N View Fig ) where it usually occurs at high elevations between around 1600 m and 1800 m, though a few specimens have been collected both lower and higher than this range. Based on collection techniques it likely inhabits leaf litter.
Comments
Despite an intensive search in the MCZC collection in collaboration with the curatorial staff, the holotype was not to be found and is presumably lost. Fortunately, however, the paratype was available for examination ( Fig. 45). In this case, we refrain from designating a neotype since the identity of D. patrizii is clear and stable. The original description does not provide any significant details that could distinguish D. patrizii from other traegaordhi - complex species, and Weber (1949) listed antennomere count (an unstable character; see Materials and Methods) and the shape of the frontal lamella as diagnostic features delineating the species from D. traegaordhi and D. hewitti . The paratype specimen however, while in less than ideal condition, is well enough preserved to associate it with recently collected material, from which a thorough description and diagnosis are possible. Despite the apparent rarity prior to this study, after association of most of the unidentified material to the type, D. patrizii turns out to be widespread in East Africa.
Variation
Varies most noticeably in color, which ranges from uniformly matte orange (as in the paratype, see Fig. 45) to dark earthy brown with yellowish appendages, as in many Tanzanian individuals ( Fig. 46 View Fig ). Pubescence is not highly variable in development, but is much less conspicuous on lighter morphs, while contrasting in darker forms. The subpetiolar process is quite variable, too, ranging from dentate with apex acute to broadly lobate, with apex rounded.








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Discothyrea patrizii Weber, 1949
| Hita-Garcia, Francisco, Lieberman, Ziv, Audisio, Tracy L., Liu, Cong & Economo, Evan P. 2019 |
Discothyrea patrizzii
| Weber 1949: 2 |
Discothyrea patrizii
| Weber 1949 |