
HADROTARSINAE
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2004.00120.x |
|
persistent identifier |
https://treatment.plazi.org/id/7E1687E1-422D-6C1E-FE9D-F9628DACFB2A |
|
treatment provided by |
Diego |
|
scientific name |
HADROTARSINAE |
| status |
|
HADROTARSINAE : CLADE 52
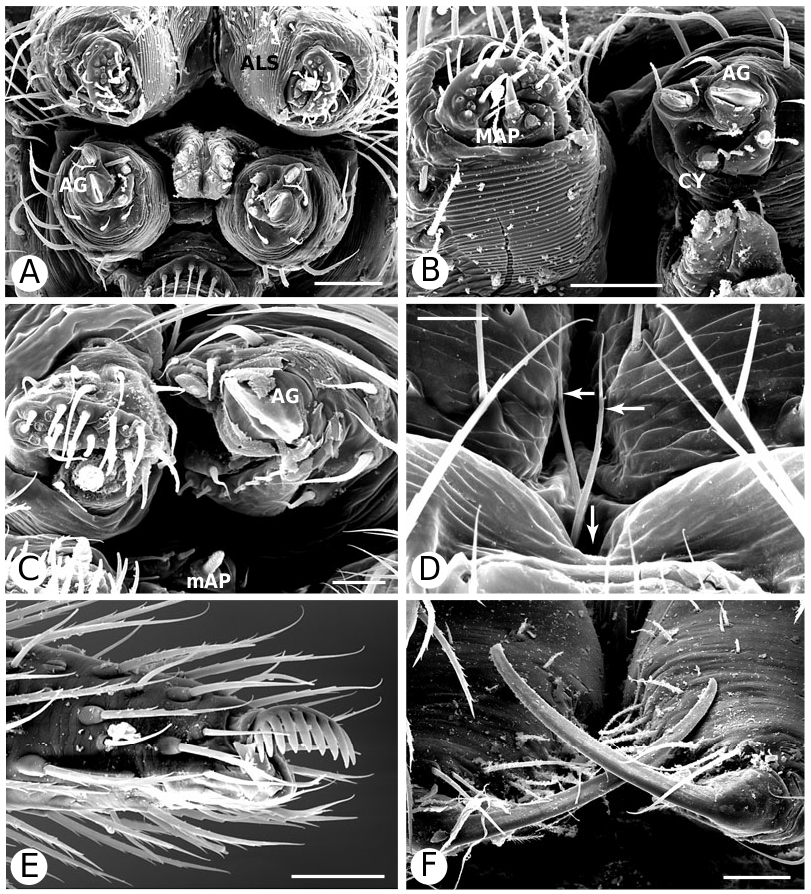
Hadrotarsine monophyly is strongly supported, although one of the putative synapomorphies, ALS stridulatory ridges, suggested by Forster et al. (1990) and Griswold et al. (1998: character 67) is doubtful. Hadrotarsus has stridulatory ridges on the ALS and Griswold et al. observed these in both Emertonella , and Dipoena . However, I have examined three Dipoena species , including their exemplar ( D. nigra Emerton ; Fig. 6D View Figure 6 ) as well as D. torva Thorell , and D. cf. hortoni (pers. observ.), and none have stridulatory ridges. Gmogala also lacks such stridulatory ridges (pers. observ.). Either Dipoena has secondarily lost this supposed hadrotarsine synapomorphy, or the character is synapomorphic for a subsidiary hadrotarsine clade [so far known only from Hadrotarsus , Emertonella , and Euryopis (203, Fig. 9B View Figure 9 )].
An identical stridulatory mechanism also occurs in Chilenodes ( Malkaridae : Palpimanoidea) ( Platnick & Forster, 1987). The placement of Chilenodes and many other palpimanoids is dubious at best ( Schütt, 2002). Classical palpimanoids are two-clawed with claw tufts, have plumose body setae, lack paracymbia and have a palpal conformation entirely different from araneoids. Chilenodes (Malkaridae) , however, along with (at least) the families Mimetidae , Holarchaeidae , Pararcheidae and Micropholcommidae (all currently Palpimanoidea, but see Schütt, 2000, 2002 for alternative placement) are three-clawed, have serrate body setae (otherwise a highly conserved araneoid synapomorphy), have a typical araneoid palpal conformation (e.g. with an embolic tegular membrane) and a paracymbium; an araneoid synapomorphy. Lehtinen (1996) came to similar conclusions based on leg cuticle ultrastructure. If the placement of Chilenodes in Araneoidea is confirmed, this character may indicate relatedness with hadrotarsines. Interestingly, another malkarid, Sternodes foraminatus Butler , shares some unique features with Phoroncidia (see Fig. 62A–D View Figure 62 , and legend).
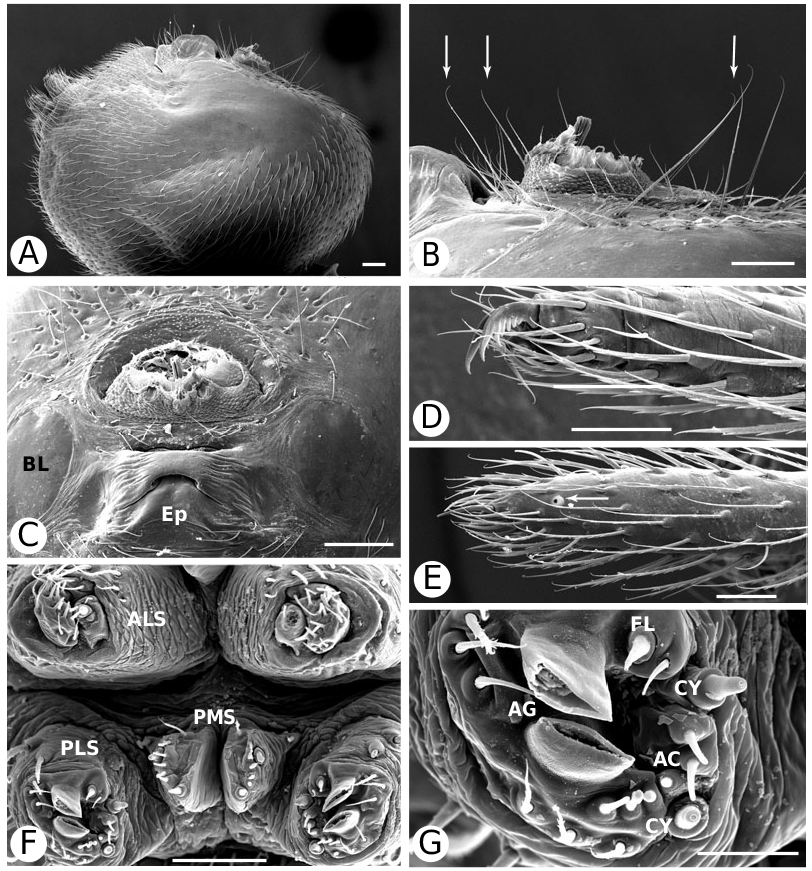
Nonetheless, several characters unite hadrotarsines including Dipoena , such as: the presence of two pairs of spermathecae (10, Fig. 93I View Figure 93 ), sperm duct entering the embolus (left palp, ventral view) counter-clockwise (60), cheliceral promargin without teeth (106, Fig. 5F View Figure 5 ), shortened paturon (117, Fig. 5C View Figure 5 ), thin cheliceral bases (118, Fig. 5D View Figure 5 ), long and slender cheliceral fangs (120, Fig. 9F View Figure 9 ), high carapace (124, Fig. 5D View Figure 5 ), carapace nearly as wide as long (126), triangular labium (136, Fig. 5A View Figure 5 ), palmate female palpal claws (177, Fig. 9E View Figure 9 ), grouped flat-tipped sensory setae on tarsus I (197, Fig. 8C, D View Figure 8 ), FL absent (212, Fig. 8A View Figure 8 ), a functional male AG (219, Fig. 6F View Figure 6 , males, as females, lack FL). The loss of prey-catching webs (224) is an ambiguous synapomorphy, but there have been no observations of prey-catching on D. nigra .
Wunderlich’s (1978) synonymy of Hadrotarsidae with Theridiidae produced a monophyletic group, but resulted in a family name with much broader circumscription and less utility. The distinctness of hadrotarsines and the absence of some classical theridiid synapomorphies - male palpal tibial rim setae (16, Fig. 13C View Figure 13 ), rim orientation (17, Fig. 10A View Figure 10 ), abdominal stridulatory picks (150, Fig. 18B–D View Figure 18 ), dorsal suprapedicellate proprioreceptors (163, Fig. 43A, B View Figure 43 ), and theridiid type tarsal comb (194, Fig. 11E, F View Figure 11 ; 195, Fig. 59A, B View Figure 59 ) - speak of the revalidation of Hadrotarsidae (see also Baert, 1984a, b), contra Wunderlich (1978) and Griswold et al. (1998). However, the phylogenetic position of this clade is contradicted in current molecular research, where Hadrotarsinae is embedded within Theridiidae ( Arnedo et al., 2004) . If correct, elevating Hadrotarsinae to family level would render Theridiidae as here circumbscribed paraphyletic; such a decision therefore awaits a combined analysis.
Griswold et al. (1998) found hadrotarsines to nest within Theridiidae , but that result is not strongly supported by their data. A single character ( Griswold et al., 1998; character 65) unifies Anelosimus and the hadrotarsines in their cladogram; colulus reduced to less than half the length of its setae. However, as discussed in this study (172–175), Anelosimus lacks a colulus, in Dipoena the colulus is clearly larger than half the length of its setae, and the condition in Emertonella (colular area strongly invaginated) is unique. If only these entries in the matrix of Griswold et al. (1998) are changed accordingly, a sister relationship between Hadrotarsinae and the remaining theridiids becomes an equally good explanation of their data.
Hadrotarsinae includes the following 14 genera: Anatea View in CoL , Audifia View in CoL , Dipoena View in CoL , Dipoenata View in CoL , Emertonella View in CoL , Euryopis View in CoL , Eurypoena Wunderlich 1992 View in CoL , Gmogala View in CoL , Guaraniella View in CoL , Hadrotarsus View in CoL , Lasaeola View in CoL , Trigonobothrys , Yaginumena View in CoL and Yoroa View in CoL .










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
HADROTARSINAE
| Agnarsson, Ingi 2004 |
Yaginumena
| Yoshida 2002 |
Eurypoena
| Wunderlich 1992 |
Dipoenata
| Wunderlich 1988 |
Guaraniella
| Baert 1984 |
Yoroa
| Baert 1984 |
Emertonella
| Bryant 1945 |
Anatea
| Berland 1927 |
Gmogala
| Keyserling 1890 |
Trigonobothrys
| , Simon 1889 |
Audifia
| Keyserling 1884 |
Hadrotarsus
| Thorell 1881 |
Lasaeola
| Simon 1881 |
Dipoena
| Thorell 1869 |
Euryopis
| Menge 1868 |