
Diarthropodella secunda, Gómez, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5051.1.12 |
|
publication LSID |
lsid:zoobank.org:pub:A99E653A-EBDF-48B1-BF24-0194136E03F9 |
|
DOI |
https://doi.org/10.5281/zenodo.5570759 |
|
persistent identifier |
https://treatment.plazi.org/id/DB1CC11E-F71F-44C6-9848-37C794CC3ABF |
|
taxon LSID |
lsid:zoobank.org:act:DB1CC11E-F71F-44C6-9848-37C794CC3ABF |
|
treatment provided by |
Plazi |
|
scientific name |
Diarthropodella secunda |
| status |
sp. nov. |
Diarthropodella secunda sp. nov.
( Figs. 36–41 View FIGURE 36 View FIGURE 37 View FIGURE 38 View FIGURE 39 View FIGURE 40 View FIGURE 41 )
urn:lsid:zoobank.org:act:DB1CC11E-F71F-44C6-9848-37C794CC3ABF
Type locality. Guaymas Basin, off Mulegé, Gulf of California , Mexico; Talud X cruise, sampling station 18 (27.1522°N, 111.6658°W); depth 1,440 m; organic carbon content, 4.14%; organic matter content, 7.13%; sand, 17.2%; clay, 11.19%; silt, 71.62 GoogleMaps %.
Specimens examined. Adult female holotype dissected and mounted onto nine slides (EMUCOP-120207-05), and one male allotype dissected and mounted onto seven slides (EMUCOP-120207-06); February 12, 2007; coll. S. Gómez.
Etymology. The specific epithet from the Latin secunda , second, makes reference to the second species attributed to the new genus. It is in the nominative singular. Gender feminine.
Description of female. Total body length measured from tip of rostrum to posterior margin of caudal rami, 405 µm; habitus pyriform, widest at posterior end of cephalothorax, tapering posteriad ( Fig. 36A View FIGURE 36 ); cephalothorax/body length ratio, 0.3.
Cephalothorax and pedigerous somites ( Fig. 36A View FIGURE 36 ) largely as in previous species.
Urosome ( Fig. 36A, C, E View FIGURE 36 ) consisting of fifth pedigerous somite (first urosomite), genital double-somite (genital—second urosomite—and third urosomite fused), two free urosomites, and anal somite. Urosomites without expansions laterally nor dorsally; integument weakly sclerotized.
Fifth pedigerous somite ( Fig. 36A View FIGURE 36 ) narrower than preceding somites; with some sensilla dorsally ( Fig. 36A View FIGURE 36 ), without spinular ornamentation.
Second and third urosomites completely fused dorsally and ventrally forming genital double-somite ( Fig. 36E View FIGURE 36 ), with dorsolateral trace of division ( Fig. 36A, C View FIGURE 36 ); genital double-somite nearly as long as wide, widest part measured in proximal third close to P6; proximal half with few dorsal sensilla and with two dorsolateral sets of spinules ( Fig. 36A, C View FIGURE 36 ), ventrally without sensilla nor spinules ( Fig. 36E View FIGURE 36 ); distal half with posterior sensilla and dorsolateral spinular rows ( Fig. 36A, C View FIGURE 36 ), ventrally with few sensilla and without spinules ( Fig. 36E View FIGURE 36 ); posterior hyaline fringe broad and smooth; genital complex hardly distinguishable, copulatory pores not exposed, paired genital apertures located ventrolaterally and covered by P6 ( Fig. 36E View FIGURE 36 ).
Fourth urosomite ( Fig. 36A, C, E View FIGURE 36 ) as distal half of genital double-somite; no pores detected.
Fifth urosomite as previous somite but without sensilla nor spinules ( Fig. 36A, C, E View FIGURE 36 ); no pores detected.
Anal somite about twice as wide as long ( Fig. 36A View FIGURE 36 ), with spinules around joint of caudal rami laterally and ventrally ( Fig. 36A, C, E View FIGURE 36 ), medial cleft without spinules ventrally ( Fig. 36E View FIGURE 36 ), with one ventral pore on each side ( Fig. 36E View FIGURE 36 ); anal operculum with minute spinules close to posterior margin, semicircular, flanked by one sensillum on each side ( Fig. 36A View FIGURE 36 ).
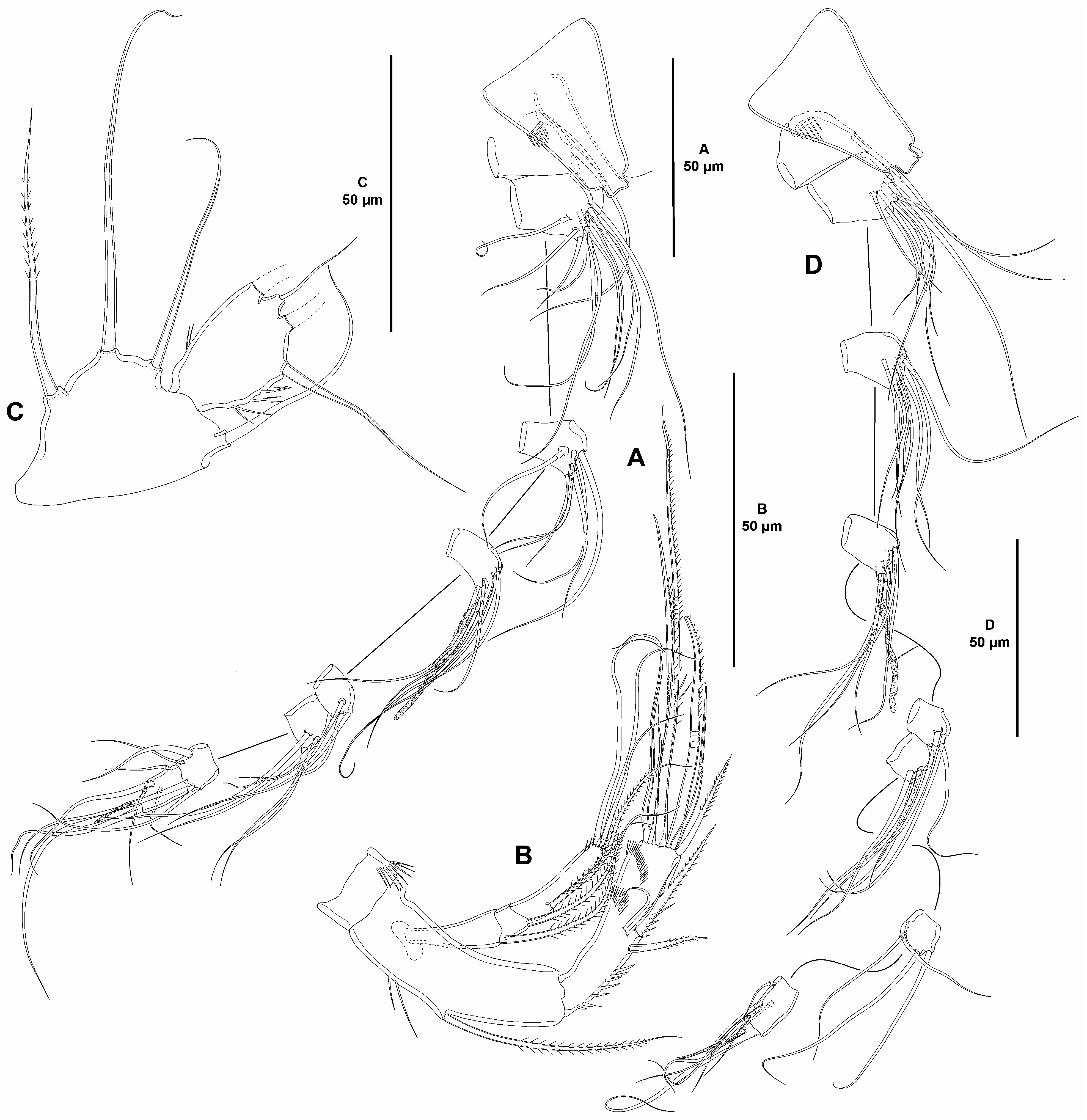
Caudal rami elongate, about four times as long as wide ( Fig. 36A View FIGURE 36 ) and as long as fifth and anal somites combined; outer and inner margins nearly straight; with outer spinules at base of setae I and II, and III, and with inner spinules subdistally ( Fig. 36A–D View FIGURE 36 ); with seven elements ( Fig. 36B, D View FIGURE 36 ); setae I and II situated subdistally, lateral, seta I very small and setiform, and ventral to seta II, the latter detached during dissection and missing in figures but its position indicated by scar; seta III subdistal, ventral ( Fig. 36B View FIGURE 36 ), detached and missing in some figures but its position indicated by scar; setae IV and seta V distal; seta VI small, issuing at inner distal corner; dorsal seta VII triarticulate at base, subdistal, arising close to inner margin.
Rostrum ( Fig. 37A View FIGURE 37 ) trapezoidal, elongate, not fused to cephalothorax, bifid, with two subdistal sensilla, without dorsal pore.
Antennule ( Fig. 37A View FIGURE 37 ) eight-segmented; all segments smooth, except for first segment with proximal spinular row; first segment without pore.All setae smooth; second and third segments with one seta with fracture plane each; seventh segment with one, eighth segment with four articulated setae. Armature formula: 1(1); 2(11); 3(7); 4(5 + (1 + ae)), 5(3); 6(4); 7(4); 8(4 + acro). Acrothek consisting of two setae and one minute aesthetasc fused basally.
Antenna ( Fig. 37B View FIGURE 37 ). Coxa short, with some outer spinules. Allobasis slightly longer than free endopodal segment; with few long inner spinules; with one abexopodal seta arising midway inner margin. Free endopodal segment elongate; with longitudinal row of spinules on proximal half and midway inner margin, without subdistal spinules, with two outer subdistal frills; armature composed of two spines and two setae laterally, distally with one inner apical spine, three apical geniculate setae and one slender element, and one outer distal strongly spinulose element fused basally to slender seta. Exopod three-segmented; first and third segments longest; first and middle segment without, third segment with spinules as shown; first and second segments with one distal seta each, third segment with one proximal and three apical setae, two of which seemingly fused basally.
Mandible ( Fig. 38A View FIGURE 38 ). Coxa relatively short. Gnathobase wide; ventral distal corner produced into small sharp semi-hyaline process; with one strong and several smaller teeth, two spines and two setae. Basis elongate, spinular ornamentation as shown, with three subdistal outer setae. Exopod arising from short pedestal, one-segmented, elongate, about 3.3 times as long as wide, and 0.4 times as long as basis, with three lateral and three apical setae, none of which fused basally. Endopod recurved, twisted over exopod, laterally with two proximal setae, one of which much longer, and one subdistal element, distally with four distal elements, of which longest fused to endopod basally and with hyaline flange in middle part.
Maxillule ( Fig. 38B View FIGURE 38 ). Arthrite of praecoxa with two surface setae and few dorsal spinules; distal armature composed of one ventral apical seta, seven apical elements, of which dorsalmost spinulose, and one lateral pinnate curved seta. Coxal endite with three setae. Basis with two endites, each seemingly with four setae. Exopod and endopod fused basally, separated from basis, one-segmented; endopod larger than exopod, with four setae; exopod with two setae.
Maxilla ( Fig. 38C–E View FIGURE 38 ). Large syncoxa with spinular ornamentation as depicted; with three endites; proximal endite bilobed, proximal lobe very small, with one seta, distal lobe with two setae; middle and distal endites elongate, the latter slightly longer, with one bare and two spinulose setae each. Basis drawn out into strong claw, with strong spine and two slender setae, one of which arising from elongate setophore. Endopod one-segmented, with six slender setae (one arising basally, two medially, and three apically).
Maxilliped ( Fig. 38F View FIGURE 38 ) subchelate. Syncoxa slightly longer than wide, nearly as long as basis; with medial spinular row midway inner margin, and with distal spinules as shown; with one bare and two spinulose strong elements, of which bare seta and one spinulose element at the same level, the other arising distally from long pedestal. Basis oval, with some outer spinules, with one anterior and one posterior inner spinular row, with two distal setae, one of which visibly longer. Endopod one-segmented, with apical claw-like element and slender seta.
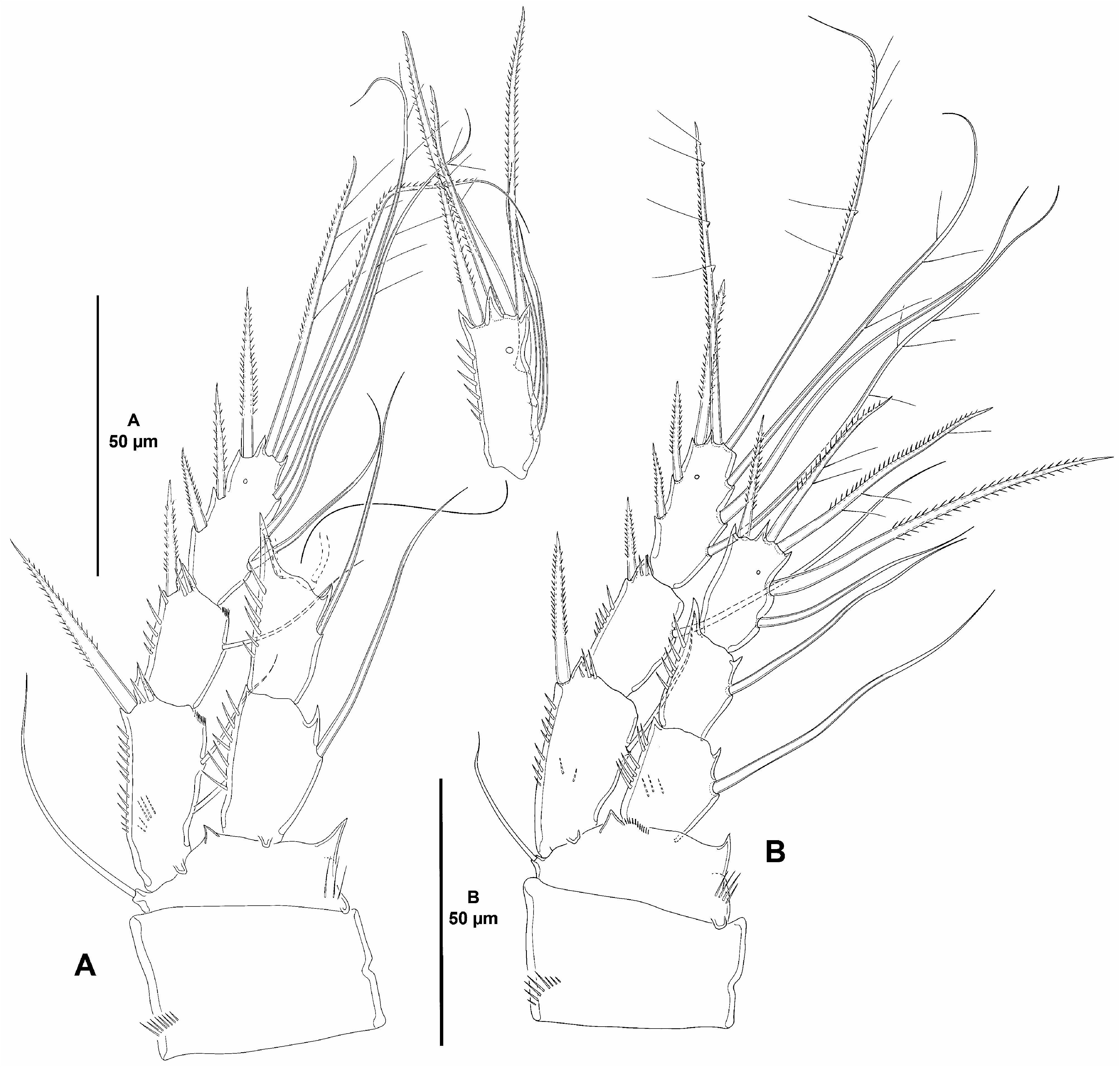
P1 ( Fig. 39A View FIGURE 39 ). Intercoxal sclerite (not shown) transversely elongate, nearly straight, without surface ornamentation. Coxa massive, 1.4 times as wide as long, with outer spinules proximally. Basis with spinules at base of outer and inner elements and between rami, with long inner proximal spinules. Exopod two-segmented, visibly shorter than endopod, reaching proximal third of ENP2; segments without outer nor inner acute distal processes; no pores detected on exopodal segments; EXP1 shorter than EXP2, with few outer spinules; EXP2 without outer ornamentation, with five elements (outermost spine probably homologous to the outer spine of the second exopodal segment of other species of the subfamily), and three elements, of which innermost long and rat-tail like in distal half, and adjacent seta short and densely setulose). Endopod two-segmented, visibly longer than exopod; ENP1 without outer nor inner outer acute distal processes, distal process of ENP2 as shown; no pores detected on endopodal segments; ENP1 reaching distal third of EXP2, 1.9 times as long as wide, and 0.8 times as long as ENP2, with longitudinal outer spinular row and with some spinules apically, with inner seta; ENP2 elongate, visibly longer than ENP1, 3.5 times as long as wide, with row of outer and apical spinules, with one inner seta issuing at proximal third, one inner apical slender seta, one medial apical pinnate element, and one outer apical spine.
P2–P4 ( Figs. 39B View FIGURE 39 , 40A–B View FIGURE 40 ). Intercoxal sclerite (not shown) as in De. prima sp. nov. Coxa and basis largely as in Diarthropodella prima sp. nov. but with minor differences in spinular ornamentation. Exopod and endopod largely as in Diarthropodella prima sp. nov.
Setal formula of swimming legs as follows:
P5 ( Fig. 37C View FIGURE 37 ). Baseoendopod pentagonal; endopodal lobe poorly-developed, with three equidistant setae separated by wide gap. Exopod oval, with some outer spinules, with four setae, of which second innermost slender.
P6 ( Fig. 37E View FIGURE 37 ) represented by a minute flap covering ventrolateral genital aperture, fused to somite, without surface ornamentation, with one slender seta.
Description of male. Total body length measured from tip of rostrum to posterior margin of caudal rami, 440 µm; general shape of habitus ( Fig. 41A View FIGURE 41 ) as in female.
Sexual dimorphism expressed in the genital somite and third urosomite not fused, P5 and P6.
Pedigerous somites largely as in female.
Genital somite and third urosomite not fused ( Fig. 41A–C View FIGURE 41 ); both somites with dorsolateral spinules and with posterior sensilla dorsally ( Fig. 41A–B View FIGURE 41 ); ventral surface of genital somite without spinular ornamentation, of third urosomite with two sets of spinules and few sensilla ( Fig. 41C View FIGURE 41 ).
Fourth urosomite with dorsolateral spinules and sensilla as shown ( Fig. 41A–B View FIGURE 41 ), ventrally with few sensilla and without spinules, no pores detected ( Fig. 41C View FIGURE 41 ).
Fifth urosomite as preceding somite but without sensilla ( Fig. 41A–C View FIGURE 41 ).
Anal somite and caudal rami ( Fig. 41A–C View FIGURE 41 ), and rostrum ( Fig. 37D View FIGURE 37 ) as in female.
Antennule ( Fig. 37D View FIGURE 37 ) as in female, not sexually dimorphic.
Antenna, mandible, maxillule, maxilla and maxilliped, P1, P2, P3, and P4 (not shown) as in female.
P5 ( Figs. 41B, C View FIGURE 41 ). Both legs fused medially forming a continuous plate; endopodal lobe poorly-developed; the left endopodal lobe possesses two seta, but the right endopodal lobe bears one seta only, being the normal condition of the armature complement uncertain; EXP small, oval, with three setae as figured.
P6 ( Figs. 41B, C View FIGURE 41 ) symmetrical, fused to somite, each leg with outer basal seta and two elements, of which innermost shortest.
Variability. No variability was detected in the female. The left endopodal lobe of the male P5 bears two setae, the right endopodal lobe bears one seta only.
s
Remarks. The basic criterion to separate the genera Stenhelia Boeck, 1865 and Delavalia was, for a long time, the segmentation pattern of the P1 ENP. The use and simplicity of this criterion, along with the subestimation of additional characters of higher significance ( Mu & Huys 2002) led many researchers to erroneous conclusions, and most stenheliins with a two-segmented P1 ENP were placed into Delavalia . Mu & Huys (2002) provided strong evidence for the monophyly of the genus Stenhelia , and challenged and abandoned the use of the segmentation pattern of P1 ENP as the only discriminant for subgeneric assignment of stenheliin taxa. The monophyly of the genus Delavalia is far from resolved, but Mu & Huys’ (2002) view was very important towards redefining the monophyly of the genus. The abandonment of the segmentation pattern of the endopod of P1 was followed by subsequent authors. Karanovic & Kim (2014) confirmed the polyphyly of Delavalia — previously suggested by Mu & Huys (2002) and Willen (2003), but see also Gómez & Cruz-Barraza (2021) —through molecular analyses and proposed three new genera with two-segmented P1 endopods, whose constituent species could have had easily been attributed to Delavalia ( Karanovic & Kim 2014) . The abandonment of the segmentation pattern of P1 ENP, the utilization of more relevant characters for generic discrimination, and the search of (syn-)apomorphies to define monophyletic groups within Delavalia are obligate steps towards the monophyly of the genus. This reasoning was followed here. The two new genera proposed here, Archaeohuysia gen. nov. and Diarthropodella gen. nov. — with two segmented P1 endopods—could have been assigned to Delavalia if the segmentation pattern of that ramus as the only generic discriminant would not had been rejected previously. These two genera were attributed to the subfamily Stenheliinae on account of the lateral displacement of the genital apertures in females, triangular and bifid rostrum, mandibular basis and endopod elongated, the latter usually with a very long seta, setation of the maxillipedal syncoxa (three setae), position of the female P5 (laterally displaced), and sexual dimorphism in the male P2 when present.
The so far monotypic genus Archaeohuysia gen. nov. resembles other stenheliin genera in most respects, but is unique in the shape, structure, and ornamentation of the endopod of P1, and probably in the imperfectly subdivided P1 EXP2 and EXP3. The modified outer spinules on P1 ENP2 forming a fan-shaped unit is hypothesized here as a potential apomorphy for the genus. The role of the latter is uncertain, but seems to work as a shovel for digging into the sediment. The retention of the primitive complement of four setae on the two-segmented P1 ENP2, and comparable subchelate non- or weakly prehensile maxillipeds are present in some other stenheliin taxa (see Gómez & Cruz-Barraza 2021), supporting their independent origin in several lineages.
Diarthropodella gen. nov. also resembles other stenheliin genera including the general structure and shape of P1 ENP and the subchelate, prehensile maxilliped.As far as I know, no other stenheliin genus displays a two-segmented P1 EXP, and the reduction from three to two segments in this ramus is regarded here as autapomorphic for the genus. Another probable apomorphy is the secondary elongation of the outer basal seta of P3. Although the male is known only for D. secunda sp. nov., the lack of sexual dimorphism in the antennule and on P2 ENP, and the fusion and symmetry of the male P6 are also potential autapomorphies for the genus. The two species presented here, Di. prima sp. nov. and Di. secunda sp. nov., are easily distinguishable by 1) the shape of the innermost setae on P1 EXP3, short, rat-tail-like and densely plumose in Di. prima sp. nov., but long and bipinnate in Di. secunda sp. nov., and 2) armature complement of the baseoendopod and exopod of the female P5 (with four and five setae, respectively, in Di. prima sp. nov., but three and four in Di. secunda sp. nov., respectively). The stenheliin species for which the males have been described exhibit sexual dimorphism in the male antennule and P2 ENP, in the relative thickness of some setae of some swimming legs in the male, division of the genital and first abdominal somite, and male P5 and P6. The significance of the lack of sexual dimorphism in the male antennule Di. secunda sp. nov. is uncertain, and the lack of sexual dimorphism in the male P2 ENP is interpreted here as a secondary loss.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Stenheliinae |
|
Genus |