
Dactylogyrus petruschewskyi Gussev, 1955
|
publication ID |
https://doi.org/10.12782/specdiv.25.349 |
|
persistent identifier |
https://treatment.plazi.org/id/03B387EE-E369-FF9C-CC89-F9FC0CD0534A |
|
treatment provided by |
Felipe |
|
scientific name |
Dactylogyrus petruschewskyi Gussev, 1955 |
| status |
|
Dactylogyrus petruschewskyi Gussev, 1955 View in CoL [New Japanese Name: Dantōbō-yubigata-mushi] ( Figs 1 View Fig , 2 View Fig )
Dactylogyrus petruschewskyi Gussev, 1955: 242–243 View in CoL , fig. 24-4; Bykhovskaya-Pavlovskaya et al. 1962: 334, fig. 663; Strelkov 1971: 72; Anonymous 1973: 132–134, figs 111– 114; Chen 1981: 151; Long and Tao 1981: 546; Gussev 1985: 161, fig. 223; Wu et al. 1991: 18, 111–112, fig. 105; Li and Zhang, 1992: 90; Jin et al. 1993a: 326, 327, fig. 95; Jin et al. 1993c: 411; Gibson et al. 1996: 25; Wang et al. 1997: 91, 116, 117; Wu et al. 2000: 24; Long 2000: 196– 197, fig. 145; Ding and Liao 2004: 629, 630, 631; Ding and Liao 2005: 246; Šimková et al. 2006: 44, 47; Gussev et al. 2010: 231, fig. 278; Singh and Chaudhary 2010: 123, 124, 125, 126; Wang et al. 2012: 45; Rajvanshi and Agrawal 2013: 580, 582; Tan 2013: 71, 133, 159, 160, 161, 192; Mendoza-Palmero et al. 2015: 4, 6; Nitta and Nagasawa 2016: 485, 486; Mendoza-Palmero et al. 2017: 154, 159; Nitta and Nagasawa 2017: 2; Šimková et al. 2017: 5, 7; Chiary and Singh 2019: 143; Daghigh Roohi et al. 2020: 1056, 1057; Nitta and Nagasawa 2020: 64.
Neodactylogyrus petruschewskyi: Yamaguti 1963: 41 , fig. 677.
Description. Body elongate, total length of body including haptor 278–291 (285, n=2), maximum width 69–97 (83, n=2) in mid-body. Cephalic region with well-developed medial and bilateral cephalic lobes. Three pairs of head organs. Head gland cells on both sides of body at level of posterior pharynx. Two pairs of eye-spots. Pharynx subspherical, 20–24 (22, n=2) long, 21–26 (24, n=2) wide; esophagus short; intestinal ceca bifurcate with caeca confluent posterior to testis.
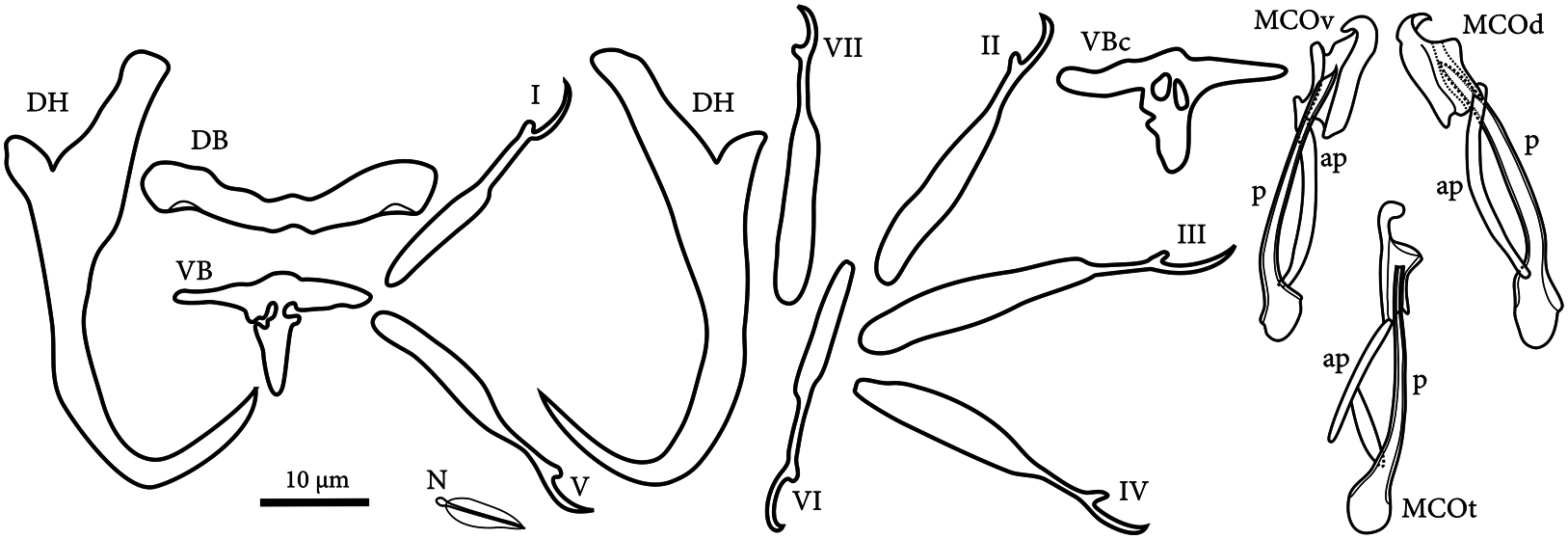
Haptor 57–61 (59, n=2) long, 67–88 (78, n=2) wide. A pair of dorsal hamulus, 35–45 (41, n=10) long; shaft slightly expanded before bent and tapering to point; length to notch 26–33 (31, n=9) long; outer root slightly tapering distally, 3–5 (4, n=9) long; inner root elongate trapezoid, 13–16 (15, n=9); point 12–15 (14, n=9) long. Dorsal bar broadly Wshaped, total length 24–30 (28, n=9), total width 7–9 (8, n=9), median width 4–5 (4, n=9). Ventral bar T-shaped, total length 20–22 (21, n=8), total width 11–14 (12, n=8); base of each side of the vertical deeply concave (n=6, Fig. 2 View Fig : VB), or concavity becoming circular holes (n=2, Fig. 2 View Fig : VBc). Marginal hooks in 7 pairs; hook length: pair I 23–27 (25, n=8); pair II 28–37 (32, n=8); pair III 30–41 (36, n=8); pair IV 28–35 (32, n=8); pair V 25–29 (27, n=8), pair VI 24–31 (28, n=8); pair VII 27–35 (32, n=8). Pair of needles 8–9 (8, n=7) long, located near the tip of fifth marginal hooks.
Gonads intercecal. Testis pyriform, posterodorsal to germarium, 32–45 (39, n=2) long, 22–23 (23, n=2) wide. Vas deferens arising from anterior margin of testis, extending from intercaecal portion, looping around left intestinal ceca onto ventral, extending seminal vesicle lying left to base of male copulatory organ, entering base of penis. Two prostatic reservoirs saccate, rounded, posterior to male copulatory organ. Male copulatory organ consisting of penis and accessory piece, 29–33 (31, n=7) long. Penis slightly curved tube, 25–29 (26, n=7) long; its base touching base of rod of accessory piece. Sclerotized accessory piece, 25–28 (26, n=7) long, consisting of half-cylindrical shaped plate with hook-like projection on sinistral tip, and rod connected to postero-dextral side of the plate; hook-like projection bending inward; plate supporting the tip of the penis dorsally; rod slightly bent (n=6), sometimes divided into two parts (n=1, Fig. 2 View Fig : MCOt).
Germarium ovate in mid-body, 47–50 (49, n=2) long, 27–30 (29, n=2) wide. Oviduct arising from anterior margin of germarium, continuing to oötype. Mehlis’ gland connecting base of oötype. Vagina unsclerotized, opening on left lateral side, mid-length of body, leading to seminal receptacle. Seminal receptacle lying left of oviduct, connecting at junction of oviduct and oötype by short, narrow duct.
Vitellaria approximately co-extensive with intestinal ceca. Egg not collected.
Materials examined. 11 specimens (NSMT-Pl 6451,
6452)
Locality. Ono River flowing into Lake Kasumigaura ( 35°58′14.5″N, 140°21′11.4″E), Inashiki City, Ibaraki Prefecture GoogleMaps .
Host. Megalobrama amblycephala Yih, 1955 ( Cypriniformes : Cyprinidae : Cultrinae ).
Site on host. Gill filaments.
Prevalence and intensity. 100% and 2 and 9 worms were collected from two fish.
Representative DNA sequences. The partial 28S rDNA (796 bp) sequences from the two specimens (NSMT-Pl 6452) were identical and submitted to the DDBJ (accession number LC538183 View Materials ) .
Etymology. The new Japanese name, “dantōbō” refers to the host, M. amblycephala , in Japanese, and “yubigatamushi” means the genus Dactylogyrus .
Remarks. Dactylogyrus petruschewskyi was originally described after being taken from the gills of Parabramis pekinensis in Lake Hanka at Astrakhanka, Primorsky Krai, Far East Russia ( Gussev 1955). The species was transferred to the genus Neodactylogyrus Price, 1938 by Yamaguti (1963), although this genus had been synonymized with Dactylogyrus Diesing, 1850 by Mizelle and Donahue (1944). As a native parasite, the monogenean was then reported with the same host species in Hubei, Zhejiang, Guangxi, and in Hunan, China ( Anonymous 1973; Wu et al. 1991; Li and Zhang 1992; Jin et al. 1993c). It was also reportedly obtained from Megalobrama terminalis (Richardson, 1846) in Hubei, Zhejiang, and in Guangxi ( Anonymous 1973; Wu et al. 1991; Li and Zhang 1992); from M. amblycephala in Hunan ( Wang et al. 2012); from Sinibrama wui (Rendahl, 1933) (Cyprinidae) as S. wui polylepis Yih and Wu, 1964 and S. wui typus Yih and Wu, 1964 in Guizhou ( Long and Tao 1981); and from Sinibrama macrops (Günther, 1868) in Zhejiang and Guangxi, China (Wu et al. 1991; Li and Zhang 1992). In addition, Gussev et al. (2010) regarded M. terminalis , listed as the parasite host by Gussev (1985), to be Megalobrama mantschuricus (Basilewsky, 1855) as M. skolkovii Dybowski, 1872 .
The measurements and morphology of sclerotized parts of the specimens collected in the present study were almost identical to descriptions of D. petruschewskyi by Gussev (1955, 1985), Anonymous (1973), Wu et al. (1991), Long (2000), and Gussev et al. (2010). There are two types of accessory piece rod: the normal type consisting of a single rod ( Gussev 1955, 1985; Gussev et al. 2010) and the tworod type ( Anonymous 1973). Both types were found in this study ( Fig. 2 View Fig ); thus, these forms are judged to demonstrate intraspecific variation.
As found through a BLAST search, the newly obtained partial 28S rDNA sequence ( LC538183 View Materials ) was identical to D. petruschewskyi ( AY548927 View Materials ). Although this accession sequence has been used for molecular phylogenetic studies on dactylogyrids ( Ding and Liao 2004, 2005; Šimková et al. 2006, 2017; Singh and Chaudhary 2010; Rajvanshi and Agrawal 2013; Tan 2013; Mendoza-Palmero et al. 2015, 2017; Nitta and Nagasawa 2016, 2017, 2020; Chiary and Singh 2019; Daghigh Roohi et al. 2020), the sequence data were not based on a published record of specimens morphologically identified. The present results support the correctness of the identification.
This dactylogyrid has been recorded in Megalobrama amblycephala from Datong Lake, Yangtze River basin, China, ( Wang et al. 2012) but the specimens have not been described morphologically nor obtained molecular information. According to morphological and molecular information, we identified the dactylogyrid taken from M. amblycephala as D. petruschewskyi for the first time in Japan. Other than D. petruschewskyi , only D. strelkowi Gussev, 1955 has been recorded from M. amblycephala ( Jin et al. 1993b) and resembles D. petruschewskyi in the T-shaped ventral bar and the shape of the dorsal hamulus ( Gussev 1955; Long 2000; Gussev et al. 2010). However, D. petruschewskyi differs from D. strelkowi which has the longer and thicker penis ( Gussev 1955; Long 2000; Gussev et al. 2010).
The internal structure of D. petruschewskyi has not been described. The results of our redescription show a common morphological feature between this species and D. bicorniculus Nitta and Nagasawa, 2016 , which is the only closely related species whose internal morphology has been described ( Nitta and Nagasawa 2016): both the prostatic reservoirs are round and small; the vagina is not sclerotized and leads parallel to the seminal receptacle; the vas deferens and the seminal vesicle do not extend above the male copulatory organ; and the testis is pyriform and located posterodorsal to the germarium. Even in recent years, the internal morphological descriptions of Dactylogyrus species are often neglected. As it consists of several genetically distinct groups ( e.g., Šimková et al. 2017), the internal morphology of Dactylogyrus will be important for future taxonomic organization of this genus, which consists of over 1,000 species ( Gibson et al. 1996; WoRMS 2020).
All recorded hosts of D. petruschewskyi are not native to Japan ( Hosoya 2015; Froese and Pauly 2019), and the dactylogyrid has not been collected in any native fish examined from the Tone River system to date (Nitta and Ishikawa, unpublished data); therefore, this species was determined to be an alien parasite that was introduced with M. amblycephala from China.
Dactylogyrus strelkowi has also been obtained from M. amblycephala ( Jin et al. 1993b) . Although we examined only two fish specimens in the present study, no invasion of D. strelkowi was found. In Datong Lake, the native habitat of M. amblycephala , D. strelkowi prevalence on this cyprinid was low, at 33% ( Jin et al. 1993b), and it has not been rediscovered in the survey by Wang et al. (2012). In contrast, all M. amblycephala examined in the area were infected by D. petruschewskyi ( Wang et al. 2012) . Based on this difference in parasitism, we conclude that only D. petruschewskyi may have been co-introduced into Japan.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Dactylogyrus petruschewskyi Gussev, 1955
| Nitta, Masato, Ishikawa, Takanori & Hagiwara, Tomiji 2020 |
Dactylogyrus petruschewskyi
| Daghigh Roohi, J. & Dalimi, A. & Pourkazemi, M. & Ghasemi, M. & Shamsi, S. 2020: 1056 |
| Nitta, M. & Nagasawa, K. 2020: 64 |
| Chiary, H. R. & Singh, H. S. 2019: 143 |
| Mendoza-Palmero, C. A. & Blasco-Costa, I. & Hernandez-Mena, D. & Perez-Ponce de Leon, G. 2017: 154 |
| Nitta, M. & Nagasawa, K. 2017: 2 |
| Simkova, A. & Benovics, M. & Rahmouni, I. & Vukic, J. 2017: 5 |
| Nitta, M. & Nagasawa, K. 2016: 485 |
| Mendoza-Palmero, C. A. & Blasco-Costa, I. & Scholz, T. 2015: 4 |
| Rajvanshi, S. & Agrawal, N. 2013: 580 |
| Tan, W. B. 2013: 71 |
| Wang, W. & Zou, W. & Tang, L. & Luo, Y. & Liu, L. & Yang, P. 2012: 45 |
| Gussev, A. V. & Gerasev, P. I. & Pugachev, O. N. 2010: 231 |
| Singh, H. S. & Chaudhary, A. 2010: 123 |
| Simkova, A. & Matejusova, I. & Cunningham, C. O. 2006: 44 |
| Ding, X. - J. & Liao, X. - H. 2005: 246 |
| Ding, X. - J. & Liao, X. - H. 2004: 629 |
| Long, S. 2000: 196 |
| Wang, W. & Li, L. & Yu, Y. & Feng, W. & Xia, C. & Wang, G. & Liu, J. & Yao, W. & Feng, S. 1997: 91 |
| Gibson, D. I. & Timofeeva, T. A. & Gerasev, P. I. 1996: 25 |
| Jin, X. & Dai, Z. & Liu, X. & Zeng, G. & Zhang, B. & He, S. & Xiang, J. 1993: 326 |
| Jin, X. & Liu, X. & Zeng, G. 1993: 411 |
| Li, G. & Zhang, J. 1992: 90 |
| Gussev, A. V. 1985: 161 |
| Chen, Z. 1981: 151 |
| Long, S. & Tao, S. 1981: 546 |
| Anonymous 1973: 132 |
| Strelkov, J. A. 1971: 72 |
| Bykhovskaya-Pavlovskaya, I. E. & Gussev, A. V. & Dubinina, M. N. & Izyumova, N. A. & Smirnova, T. S. & Sokolovskaya, I. L. & Shtein, G. A. & Shul'man, S. S. & Epshtein, V. M. 1962: 334 |
| Gussev, A. V. 1955: 243 |