Cyrtodactylus thuongae
sp. nov.
Figs. 1
View FIGURE 1
, 2
View FIGURE 2
A, 3
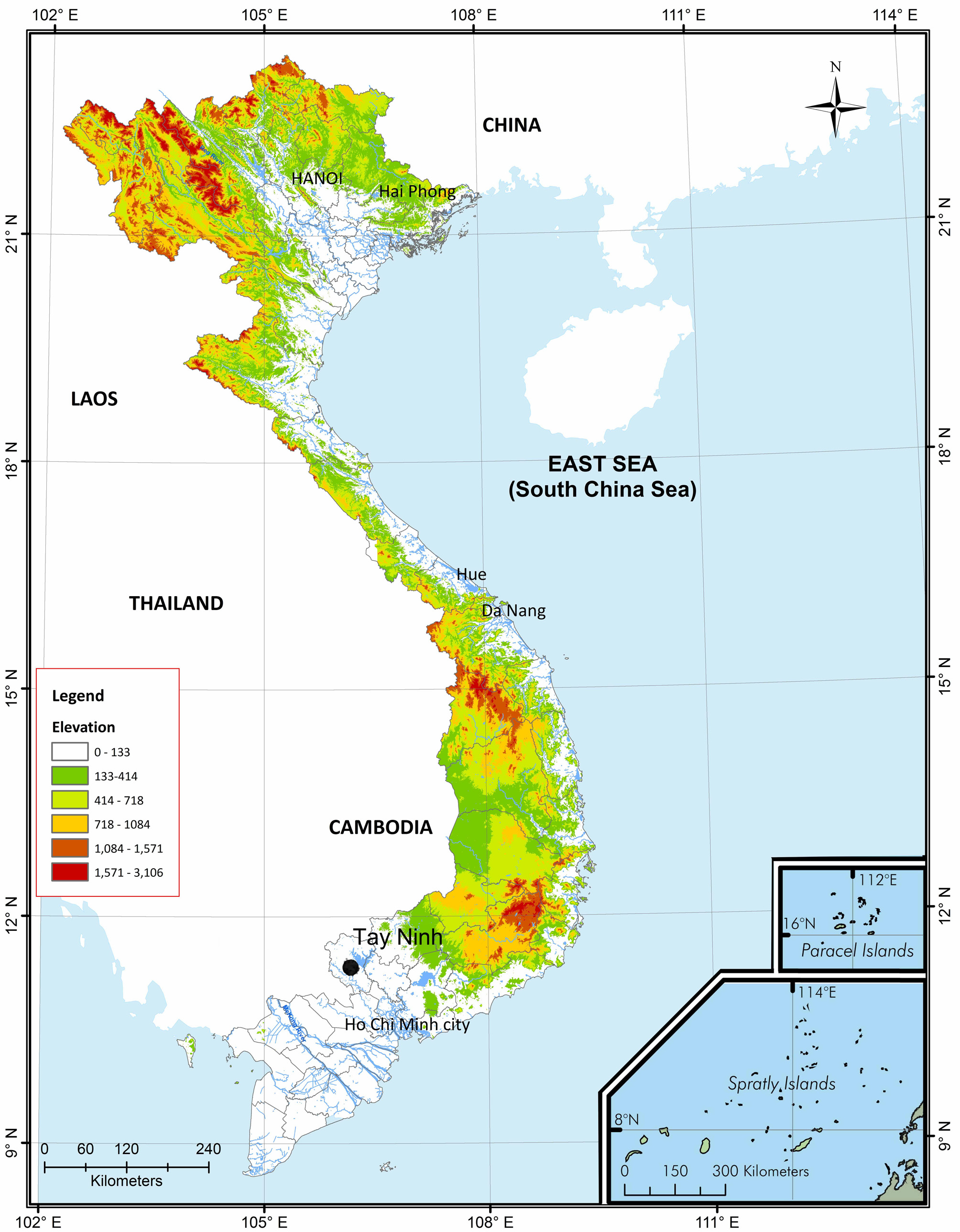
Holotype. IEBR A.2013.23, adult male, collected on 14 May 2013 by T.M. Phung in a rocky cave from Ba Den Mountain, Thanh Tan Ward, Tay Ninh Town, Tay Ninh Province ( 11°21'52"N 106°10'27"E), southern Vietnam, at an elevation of 600 m a.s.l.
Paratypes. IEBR A.2013.24, adult female, IEBR A.2013.25, adult female, VNMN A.2013.12, adult female, were collected on 16 December 2012; VNMN A.2013.13, adult male, ZFMK 94994–94997, adult females, and ZFMK 94998–94999, subadults, were collected on 14 May 2013 by T.M. Phung, the same collection data as the holotype.
Diagnosis. A medium-sized
Cyrtodactylus
with a maximum SVL of 77.6 mm, distinguished from all congeners by a combination of the following characters: 1) dorsal pattern consisting of irregular blotches on dorsum and occiput; nuchal loop sometimes interrupted medially; 2) precloacal pore absent or one in males; 3) femoral pore series separated from precloacal pore series, 0–3 on each thigh in males; 4) ventrals in 29–44 longitudinal rows at midbody; 5) lateral skin folds present, without interspersed tubercles; 6) dorsal tubercles in 16–18 irregular rows; 7) enlarged femoral scales and precloacal scales present; 8) postcloacal spurs two or three; 9) subcaudal scales slightly enlarged; and 10) precloacal groove absent.
Description of holotype. Size small (SVL 67.6 mm), tail regenerated (TaL 41.55 mm); supralabials 8/9; infralabials 7/8; nares bordered by rostral anteriorly, first supralabial laterally and four or five nasals posteriorly; supranasals separated from each other by a pentagonal internasal and two postnasals; upper anterior ciliaries three times larger than posterior ciliaries; dorsal scales of head granular; mental square, as wide as rostral; one pair of enlarged postmentals, longer than wide, in contact with each other posteriorly about a half of their length, bordering mental anteriorly, first two labials laterally, and six gular scales posteriorly; outer pair of gular scales is as large as postmentals; dorsal scales somewhat granular to flattened; dorsal tubercles round, conical, surrounded by 10 granular scales, in 17 irregular longitudinal rows at midbody; ventral scales smooth, in 36 longitudinal rows at midbody; lateral folds present, poorly developed, without interspersed tubercles; upper and lower surface of fore limb without distinct tubercles; dorsal surface of hind limb covered by distinctly developed, round, conical tubercles; fingers and toes lacking distinct webbing; lamellae under fourth finger 15/14 and under fourth toe 16/17; claws surrounded by a small scale on upper and a large scale on lower sides; series of distinctly enlarged femoral scales separated from enlarged precloacal scales by 15 or 16 small, poreless femoral scales; femoral pores absent; precloacal pore one; enlarged precloacal scales nine, including one pore-bearing scale; precloacal region covered by a patch of approximately 23 enlarged scales behind precloacal pore-bearing scale; precloacal groove absent; postcloacal tubercles 1/2, enlarged, on lateral surface of slight hemipenial swelling; hemipenes everted; tail regenerated, dorsum of original tail bearing distinct round tubercles; subcaudals not distinctly enlarged.
Coloration in ethanol. Ground coloration light grey, with dark brown dorsal pattern; dorsum with dark blotches, lighter on the sides; legs, including digits and toes and dorsal surface of tail lighter with dark brown blotches and irregular faint brownish markings between blotches; dorsal surface of head grey with dark irregular blotches on the occiput, snout slightly spotted; venter light cream; ventral surface of tail grey; tubercles on edges of blotches lighter in color; nuchal loop distinct, interrupted and tapering medially. For coloration in life see Fig. 1
View FIGURE 1
A.
Variation of paratypes. Two subadults (ZFMK 94998–94999) are uniformly light grey in ground colouration and have distinct brown blotches on dorsum and occiput; limbs and snout are uniformly light grey and their nuchal loops are distinct, complete and tapering medially. The male paratype VNMN A.2013.13 lacks precloacal and femoral pores. Internasal scales vary from two (in the paratype VNMN A.2013.13) to three (in the paratypes ZFMK 94994, 94995 and 94997). For further variation in scalation see Table 1, and for colour pattern variation see Figure 3
View FIGURE 3
.
Comparisons. Comparisons are based on the original descriptions or descriptions provided in broader faunal and taxonomic publications (e.g., Grismer et al. 2008; Rösler & Glaw 2008; Bauer et al. 2009, 2010; Ngo & Grismer 2010; Ngo & Pauwels 2010; Sumontha et al. 2010; Ziegler et al. 2010; David et al. 2011; Iskandar et al. 2011; Luu et al. 2011; Ngo 2011; Ngo & Chan 2011; Schneider et al. 2011; Nazarov et al. 2012; Ngo & Grismer 2012, Ziegler et al. 2013).
Cyrtodactylus thuongae
sp. nov. differs from its Vietnamese congeners by the following characters (see Table 2):
Cyrtodactylus thuongae
sp. nov. has no or one precloacal pore, not in a continuous series with 0–3 femoral pores on each side in males and thus differs from:
C. bichnganae Ngo & Grismer 2010
(8–10 PP),
C. bidoupimontis Nazarov, Poyarkov, Orlov, Phung, Nguyen, Hoang & Ziegler 2012
(4–6 PP),
C. bugiamapensis Nazarov, Poyarkov, Orlov, Phung, Nguyen, Hoang & Ziegler 2012
(7 or 8 PP),
C. caovansungi Orlov, Nguyen, Nazarov, Ananjeva & Nguyen, 2007
(9 PP),
C. cattienensis Geissler, Nazarov, Orlov, Böhme, Phung, Nguyen & Ziegler, 2009
(6–8 PP),
C. chauquangensis Hoang, Orlov, Ananjeva, Johns, Hoang & Dau, 2007
(6 or 7 PP),
C. condorensis Smith 1921a
(4–7 PP),
C. cryptus Heidrich, Rösler, Vu, Böhme & Ziegler 2007
(9–11 PP),
C. hontreensis Ngo, Grismer & Grismer 2008
(7 or 8 PP),
C. huongsonensis Luu, Nguyen, Do & Ziegler 2011
(6 PP),
C. huynhi Ngo & Bauer, 2008
(7–9 PP),
C. intermedius Smith
(8–10 PP),
C. irregularis Smith 1921b
(5–7 PP),
C. kingsadai Ziegler, Phung, Le & Nguyen 2013
(7–9 PP),
C. martini Ngo 2011
(4 PP),
C. phongnhakebangensis Ziegler, Rösler, Herrmann & Thanh 2002
(32–42 PP+FP),
C. phuocbinhensis Nguyen, Le, Tran, Orlov, Lathrop, Macculloch, Le, Jin, Nguyen, Nguyen, Hoang, Che, Murphy & Zhang, 2013
(7 PP),
C. phuquocensis Ngo, Grismer & Grismer, 2010
(7–9 PP),
C. pseudoquadrivirgatus Rösler, Vu, Nguyen, Ngo & Ziegler, 2008
(5–9 PP), and
C. roesleri Ziegler, Nazarov, Orlov, Nguyen, Vu, Dang, Dinh & Schmitz 2010
(20–28 PP+FP),
C. takouensis Ngo & Bauer 2008
(3 or 4 PP),
C.taynguyenensis Nguyen, Le, Tran, Orlov, Lathrop, Macculloch, Le, Jin, Nguyen, Nguyen, Hoang, Che, Murphy & Zhang, 2013
(6 PP),
C. thochuensis Ngo & Grismer 2012
(3–5 PP),
C. yangbayensis Ngo & Chan, 2010
(6–8 PP) and
C. ziegleri Nazarov, Orlov, Nguyen & Ho, 2008
(5–8 PP).
Cyrtodactylus thuongae
sp. nov. differs from
C. badenensis Nguyen, Orlov & Darevsky 2006
,
C. grismeri Ngo 2008
and
C. nigriocularis Nguyen, Orlov & Darevsky 2006
by having enlarged femoral scales and lacking transversely enlarged subcaudals.
Cyrtodactylus thuongae
sp. nov. differs from
C. cucphuongensis Ngo & Chan 2011
, by the absence of transversely enlarged subcaudals, having fewer enlarged femoral scales (2–4 vs. 14) and the presence of femoral and precloacal pores in males.
Cyrtodactylus thuongae
sp. nov. differs from
C. eisenmanae Ngo, 2008
by having more dorsal tubercle rows (16–18 vs. 14), fewer ventral scale rows (29–44 vs. 44 or 45), the lack of transversely enlarged subcaudals, and the presence of femoral and precloacal pores in males.
Cyrtodactylus thuongae
sp. nov. differs from
C. paradoxus ( Darevsky & Szczerbak 1997)
by the presence of femoral pores in males, the absence of distinctly enlarged subcaudal scales and a different body pattern (blotched vs. banded).
With respect to the remaining
Cyrtodactylus
(except for zoogeographically distant species from the Papua- Australian region, the Solomon Islands, India, Sri Lanka, the Nicobar islands and Nepal),
Cyrtodactylus thuongae
sp. nov. has no or one precloacal pore (PP), separated from a series of 0–3 femoral pores (FP) on each side in males (0 or 1+0–3 PP+FP) and thus differs from the following species that have more precloacal and femoral pores:
C. agusanensis ( Taylor 1915)
(7–11+3–14),
C. astrum Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels, 2012
(31–38 PP and FP in a continuous series),
C. auribalteatus Sumontha, Panitvong & Deein, 2010
(6+4–5),
C. australotitiwangsaensis Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(39–45),
C. ayeyarwadyensis Bauer 2003
(10–28),
C. batucolus Grismer, Chan, Grismer, Wood & Belabut 2008
(43–46),
C. bintangrendah Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(41–46),
C. bintangtinggi Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(38–41),
C. brevipalmatus (Smith)
(7–10+6–7),
C. chanhomeae Bauer, Sumontha & Pauwels 2003
(32–34),
C. consobrinoides ( Annandale 1905)
(26),
C. deveti ( Brongersma 1948)
(18–22),
C. dumnuii Bauer, Kunya, Sumontha, Niyomwan, Pauwels, Chanhome & Kunya 2010
(5–6+6),
C. feae ( Boulenger 1893)
(32),
C. fumosus (Müller 1995)
,
C. gansi Bauer 2003
(42–52),
C. halmahericus Mertens 1929
(42–52),
C. jarujini Ulber 1991
(42– 54),
C. langkawiensis Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(30),
C. lekaguli Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(30–36),
C. lomyenensis Ngo & Pauwels 2010
(32–40),
C. macrotuberculatus Grismer & Norhayati 2008
(28–40),
C. phuketensis Sumontha, Pauwels, Kunya, Nitikul, Samphantamit & Grismer, 2012
(33– 36),
C. pulchellus Gray 1827
(33–39),
C. redimiculus King 1962
(5–8+8–9),
C. seribuatensis Youmans & Grismer 2006
(40–44),
C. surin
Chan-ard & Makchai, 2011 (34),
C. tamaiensis Mahony 2009
(40),
C. trilatofasciatus Grismer, Wood, Quah, Anuar, Muin, Sumontha, Ahmad, Bauer, Wangkulangkul, Grismer & Pauwels 2012
(41– 46),
C. variegatus ( Blyth 1861)
(32) and
C. wetariensis ( Dunn 1927)
(11–12+12–13).
Cyrtodactylus thuongae
sp. nov. has a series of 0–3 femoral pores on each side and no or one precloacal pore in males and thus differs from the following species which lack femoral pores and have higher precloacal pore counts:
C. aequalis Bauer 2003
(9 PP),
C. annandalei Bauer 2003
(11–12),
C. annulatus ( Taylor 1915)
(6),
C. baluensis ( Mocquard 1890)
(9–11),
C. chrysopylos Bauer 2003
, (10),
C. consobrinus ( Peters 1871)
(9–11),
C. durio Grismer, Anuar, Quah, Muin, Onn, Grismer & Ahmad 2010
(12),
C. elok Dring 1979
(7 or 8),
C. erythrops Bauer, Kunya, Sumontha, Niyomwan, Panitvong, Pauwels, Chanhome, & Kunya, 2009
(9),
C. ingeri Hikida 1990
(8),
C. interdigitalis Ulber 1993
(14),
C. khasiensis ( Jerdon 1870)
(12–14),
C. majulah Grismer, Wood & Lim, 2012
(7–11),
C. mandalayensis Mahony 2009
(5),
C. mimikanus ( Boulenger 1914)
(7–17),
C. nuaulu Oliver, Edgar, Mumpuni, Iskandar & Lilley 2009
(6),
C. payacola Johnson, Quah, Anuar, Muin, Wood, Grismer, Greer, Chan, Ahmad, Bauer & Grismer, 2012
(11 or 12),
C. pantiensis Grismer, Chan, Grismer, Wood & Belabut 2008
(8 or 9),
C. peguensis ( Boulenger 1893)
(7–9),
C. russelli Bauer 2003
(15),
C. slowinskii Bauer 2002
(9–11),
C. spinosus Linkem, McGuire, Hayden, Setiadi, Bickford & Brown 2008
(12–13),
C. sworderi ( Smith 1925)
(8 or 9),
C. teyniei David, Nguyen, Schneider & Ziegler 2011
( 14 in the female holotype),
C. tigroides Bauer, Sumontha & Pauwels 2003
(7 or 9)
C. wakeorum Bauer 2003
(12) and
C. wayakonei Nguyen, Kingsada, Rösler, Auer & Ziegler 2010
(6– 8).
Cyrtodactylus thuongae
sp. nov. differs from the following
Cyrtodactylus
species by the absence of precloacal and femoral pores in females:
C. batik Iskandar, Rachmansah & Umilaela 2011
,
C. darmandvillei ( Weber 1890)
,
C. gordongekkoi ( Das 1994)
,
C. jarakensis Grismer, Chan, Grismer, Wood & Belabut 2008
,
C. laevigatus ( Darevsky 1964)
,
C. thirakhupti Pauwels, Bauer, Sumontha & Chanhome 2004
and
C. wallacei Hayden, Brown, Gillespie, Setiadi, Linkem, Iskandar, Umilaela, Bickford, Riyanto, Mumpuni & McGuire 2008
.
Cyrtodactylus thuongae
sp. nov. has 29–44 ventral scale rows at midbody and thus differs from
C. agamensis ( Rösler & Glaw 2008)
(67),
C. gubaot Welton, Siler, Linkem, Diesmos & Brown, 2010
(54–67),
C. lateralis (Werner 1986)
(60–64),
C. malayanus ( De Rooij 1915)
(58–62),
C. mamanwa Welton, Siler, Linkem, Diesmos & Brown, 2010
(57–70),
C. matsuii Hikida 1990
(51),
C. sanook Pauwels, Sumontha, Latinne & Grismer, 2013
(27– 28), and C. s umuroi Welton, Siler, Linkem, Diesmos & Brown 2010 (53–58).
Cyrtodactylus thuongae
sp. nov. has 16–18 dorsal longitudinal tubercle rows and thus differs from
C. brevidactylus Bauer 2002
(27),
C. buchardi David, Teynié & Ohler 2005
(25),
C. oldhami ( Theobald 1876)
(19– 24) which lack enlarged femoral scales,
C. redimiculus King 1962
(14–16), which lacks a ventro-lateral fold,
C. sumonthai Bauer, Pauwels & Chanhome 2002
(12) and
C. quadrivirgatus Taylor 1962
, which lack femoral pores.
Cyrtodactylus thuongae
sp. nov. differs from
C. pageli Schneider, Nguyen, Schmitz, Kingsada, Auer & Ziegler 2011
by lacking precloacal pores in females (vs. 5 or 6).
Cyrtodactylus thuongae
sp. nov. differs from
C. aurensis Grismer 2005
,
C. cavernicolus Inger & King 1961
,
C. jambangan Welton, Siler, Diesmos & Brown 2010
,
C. jellesmae (Boulenger 1887)
,
C. marmoratus Gray 1831
,
C. papilionoides Ulber & Grossmann 1991
,
C. philippinicus ( Steindachner 1867)
,
C. pubisulcus Inger 1958
C. rubidus ( Blyth 1861)
,
C. semenanjungensis Grismer & Leong 2005
C. stresemanni Rösler & Glaw 2008
,
C. tautbatorum Welton, Siler, Diesmos & Brown 2009
,
C. tiomanensis Das & Lim 2000
, by the absence of a precloacal groove.
Cyrtodactylus thuongae
sp. nov. differs from
C. yoshii Hikida 1990
and
C. zhaoermii Shi & Zhao 2010
by the presence of femoral pores in males. The new species further differs from
C. zhaoermii
by having fewer enlarged femoral scales (2–4 vs.12–14).
Cyrtodactylus thuongae
sp. n. differs from
C. leegrismeri Chan & Norhayati 2010
by the absence of enlarged subcaudals, having fewer precloacal pores in males (absent or one vs. six) and the presence of femoral pores in males.
Cyrtodactylus thuongae
sp. n. differs from
C. angularis ( Smith 1921b)
by the presence of enlarged femoral scales and by having fewer precloacal pores in males (absent or one vs. six).
Distribution.
Cyrtodactylus thuongae
is currently known only from the type locality in Tay Ninh Province, Vietnam (see Fig. 5
View FIGURE 5
).
Etymology. The species name is dedicated to the wife of the first author, Dr. Thuong Thi Lien Nguyen from University of Food Industry, Ho Chi Minh City, Vietnam, for support of his field research in Vietnam.
Natural history notes.
Cyrtodactylus thuongae
inhabits the same habitat with two other co-occurring cave dwelling species,
C. badenensis Nguyen, Orlov & Darevsky, 2006
and
C. nigriocularis Nguyen, Orlov & Darevsky, 2006
( Fig. 2
View FIGURE 2
). Specimens were collected at night (ca. 22–24:00) on the rocky wall in a small cave, approximately 0.5–2.5 m above the ground. Most of the specimens were found inside the cave (ca. 2–5 m from the entrance) and only one was collected outside (ca. 1.5 m away from the entrance). The surrounding habitat was mixed forest of hardwood and vine, dominated by trees of the families
Moraceae
,
Fabaceae
, and
Dipterocarpaceae
.
C. badenensis
and
C. nigriocularis
were also found on rocky cliffs of the cave, usually at heights below 1.5 m above the ground.