
Cyanocohniella calida Kaštovský, Berrendero, Hladil et Johansen, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.181.5.3 |
|
persistent identifier |
https://treatment.plazi.org/id/333C87D8-8179-DD6C-FF21-FF75FD88FBEF |
|
treatment provided by |
Felipe |
|
scientific name |
Cyanocohniella calida Kaštovský, Berrendero, Hladil et Johansen |
| status |
sp. nov. |
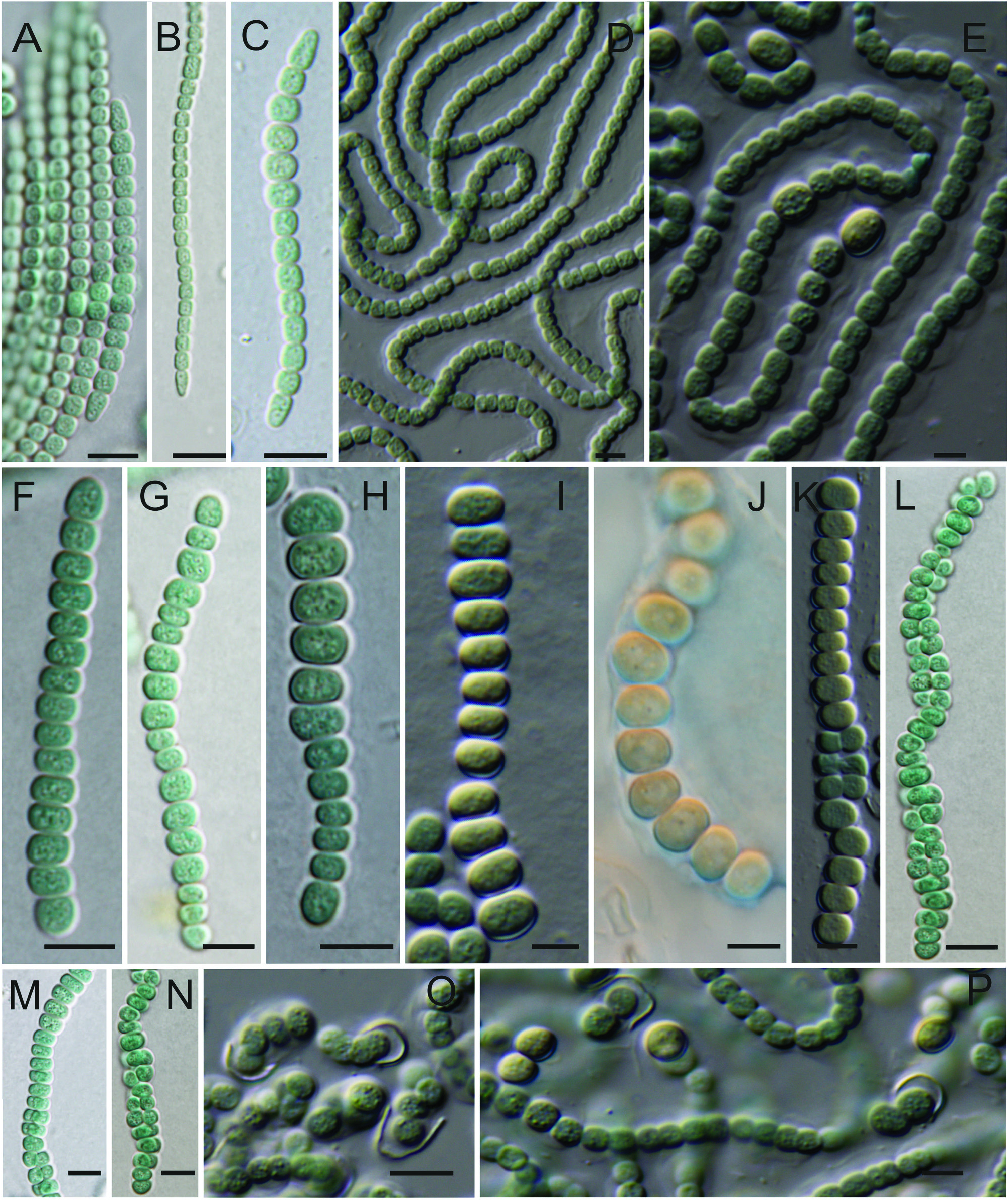
Cyanocohniella calida Kaštovský, Berrendero, Hladil et Johansen , sp. nov. ( Fig. 2 View FIGURE 2 ).
Type:— CZECH REPUBLIC. Karlovy Vary, 50°13’06.0”N, 12°53’08.6”E 445 m a.s.l., J. Hladil, 2012 (holotype CBTS! A-023 , Herbarium for Nonvascular Cryptogams at the Department of Botany , Faculty of Science , University of South Bohemia, Czech Republic ) GoogleMaps . Reference Strain: Cyanocohniella calida CCALA 1049 (Culture Collection of Autotrophic Organisms at the Institute of Botany , Třeboň, CZ) .
Etymology:—’ calida’ (L.) = warm or hot, named for its thermotolerance.
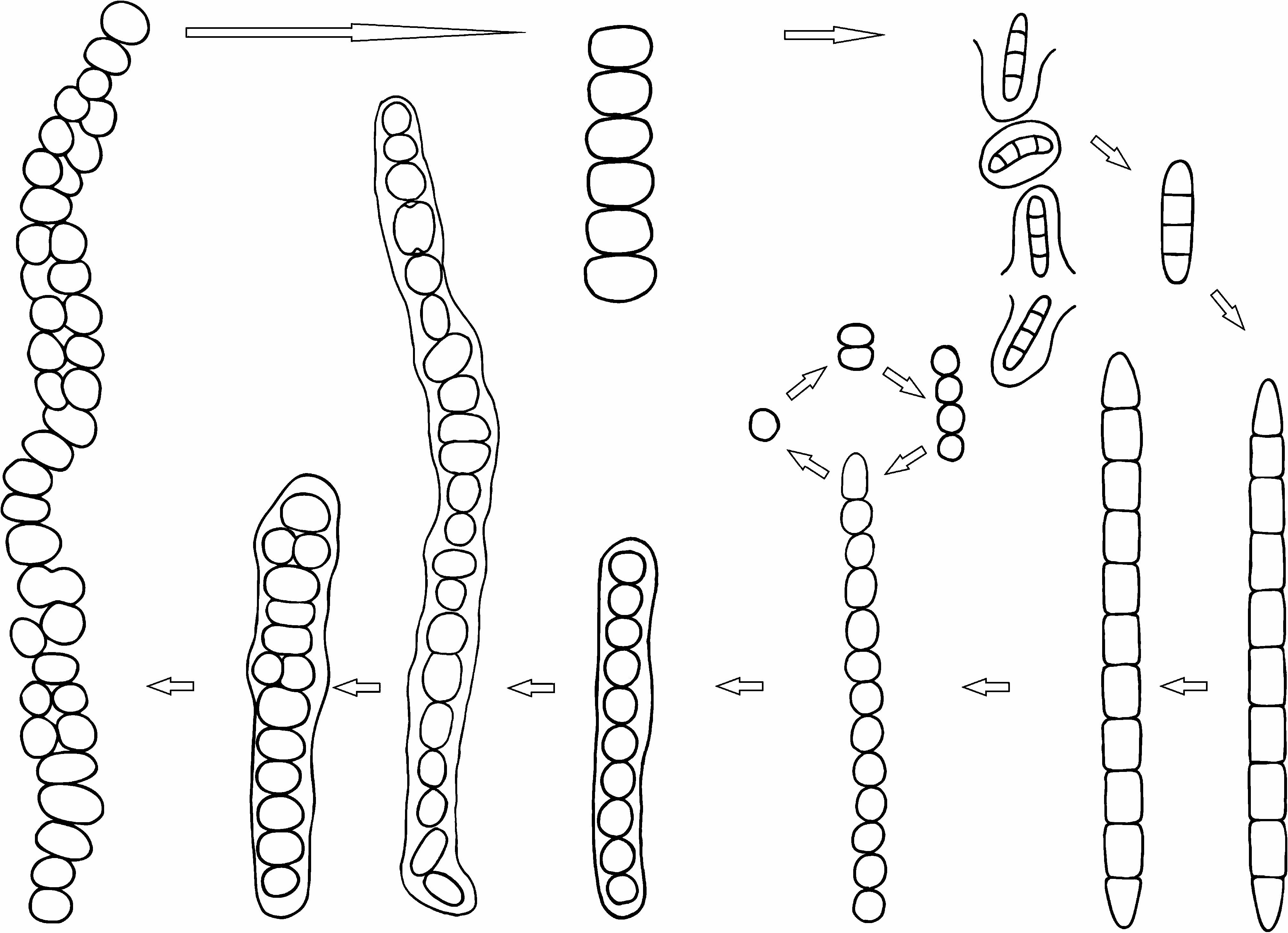
Observations:— Without examination of the entire life cycle, the stages of Cyanocohniella are similar to several previously described cyanobacterial taxa.Young filaments resemble Leptolyngbya Anagnostidis & Komárek (1988: 390) or Pseudanabaena Lauterborn (1915: 437) . For the majority of its life cycle filaments resemble Nostoc Vaucher ex Bornet & Flahault (1888: 181) , while the late stage biseriate filaments with irregular cells are similar to Chlorogloeopsis Mitra & Pandey (1967: 111) . At times, different life cycle stages are evident in single filaments (see Figs. 1C and H View FIGURE 1 ). It is possible that Hapalosiphon laminosus f. nostocoides and H. laminosus var. oscillarioides Frémy (1936: 182) are conspecific with C. calida . However, we list these only as possible synonyms. They are taxa at a different rank than species so have no nomenclatural standing in the naming of this taxon (Art. 11.4 in McNeill et al. 2012). They are poorly known taxa from African hot springs, and even if material were available, it would be nearly impossible to say if they were or were not identical to the Karlovy Vary material based on morphology alone. We recommend that these taxa be considered in need of study and revision, and if they are again recovered from African warm springs molecular characterization could determine if they belong to Cyanocohniella or some other genus. In natural conditions this species lives in hot water or atmophytically close to the hot spring. The type locality is periodically flooded with cold stream water, and populations must re-establish after these events. It consequently appears that the taxon is thermotolerant rather than obligately stenothermal. In culture, the species does not require elevated temperatures and grows without difficulty at room temperature. We hypothesize that the thermotolerance allows this taxon to escape competition with the Pseudanabaenales and Oscillatoriales that grow in thick mats on the periphery of the springs.
Evidence of phylogenetic separation:— Cyanocohniella is very distant from the thermal Mastigocladus laminosus strains that have been sequenced ( Fig. 3 View FIGURE 3 ), and in our larger phylogeny still places sister to several genera in the Aphanizomenonaceae . The newly isolated strain CCALA 1049 clusters tightly with strain Kaštovský 1996/2, and both strains are certainly conspecific. In our phylogeny, Cyanocohniella was resolved as a sister taxon to the clade containing Anabaenopsis Miller (1923: 125) and Cyanospira Florenzano et al. (1985: 305) , with bootstrap support in the maximum likelihood and probability in Bayesian analysis; bootstrap of maximum parsimony is only 60 ( Fig. 3 View FIGURE 3 ). It was also closely related to Nodularia Mertens ex Bornet & Flahault (1888: 243) and Chrysosporum Zapomělová et al. (2012: 359) , with similar support. The remainder of the genera in the Aphanizomenonaceae was not resolved with bootstrap support with regards to their relationship with the in-group taxa containing Cyanocohniella , or even among other taxa in the Nostocales . The Aphanizomenonaceae is problematic because 16S rRNA sequence similarity exceeds 97.6% for all taxa in the family, which contains very morphologically and biochemically diverse species and genera. Most members of the family are capable of producing aerotopes. However, some benthic taxa (certain Nodularia species , Anabaena Bory de Saint-Vincent ex Bornet & Flahault (1888: 180 , 224) sensu stricto, and Trichormus variabilis (Kützing ex Bornet & Flahault 1888: 226) Komárek & Anagnostidis (1989: 304) lack aerotopes. Cyanocohniella lacks aerotopes, and this is evidence of its evolutionary separation from the other members of its clade which are obligately planktonic ( Fig. 3 View FIGURE 3 ). It is further separated from Anabaenopsis and Cyanospira by the complete absence of spiraling trichomes. The Mastigocladus strains demonstrating true branching were all contained in a clade of Fischerellaceae / Mastigocladaceae / Hapalosiphonaceae distant from the Aphanizomenonaceae . The strains of M. laminosus from thermal habitats were grouped in the same clade together with exclusively thermal strains ascribed to Fischerella . Fischerella indica CALU 987 ( EU116033 View Materials ) from soil grouped with Fischerella sp. CENA 19 ( AY039703 View Materials ), a strain also isolated from soil. There was a complete phylogenetic divide between the thermal true-branching strains and the soil-inhabiting true-branching strains. Cyanocohniella was very distant from all true-branching taxa.According to the comparison of the 16S rRNA sequence similarities, the two strains of Cyanocohniella calida had a similarityof 99.9% ( Table 1). Cyanospira ripkkae ( FR774774 View Materials ), Anabaenopsis elenkinii Miller (1923: 125) ( AM773308 View Materials ) and Nodularia harveyana Thuret ex Bornet et Flahault (1888: 243) ( AF268019 View Materials ) were the most similar taxa to C. calida (98%, 97.1% and 97.2%, respectively). The Mastigocladu s/ Fischerella thermal strains clade had very low similarities of 89.8–90.5% with C. calida CCALA 1049 . Fischerella indica CALU 987 and Fischerella sp. CENA19 were also more distantly related, with similarity values of 92.0 and 91.2% to C. calida , respectively ( Table 1).
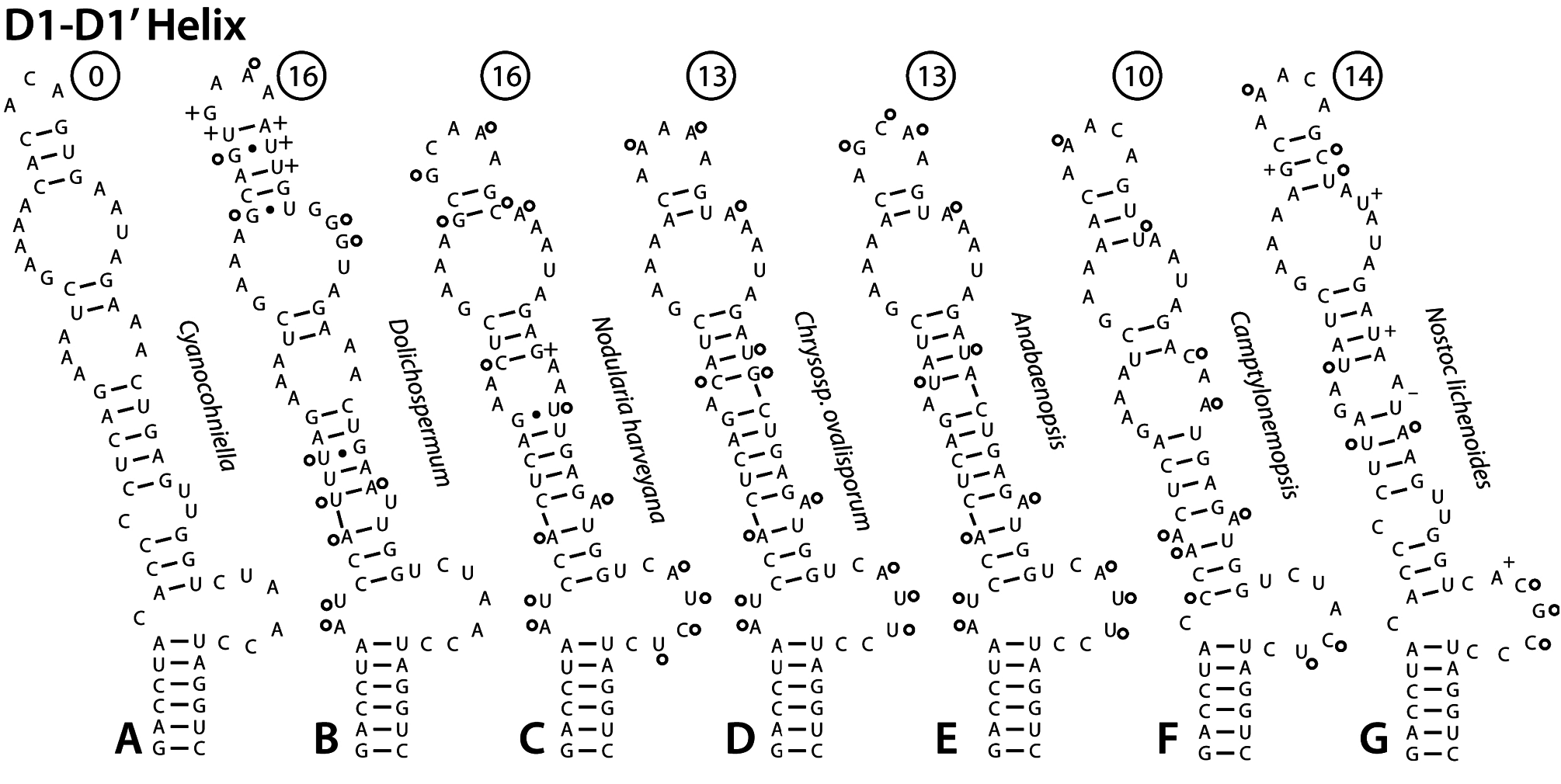
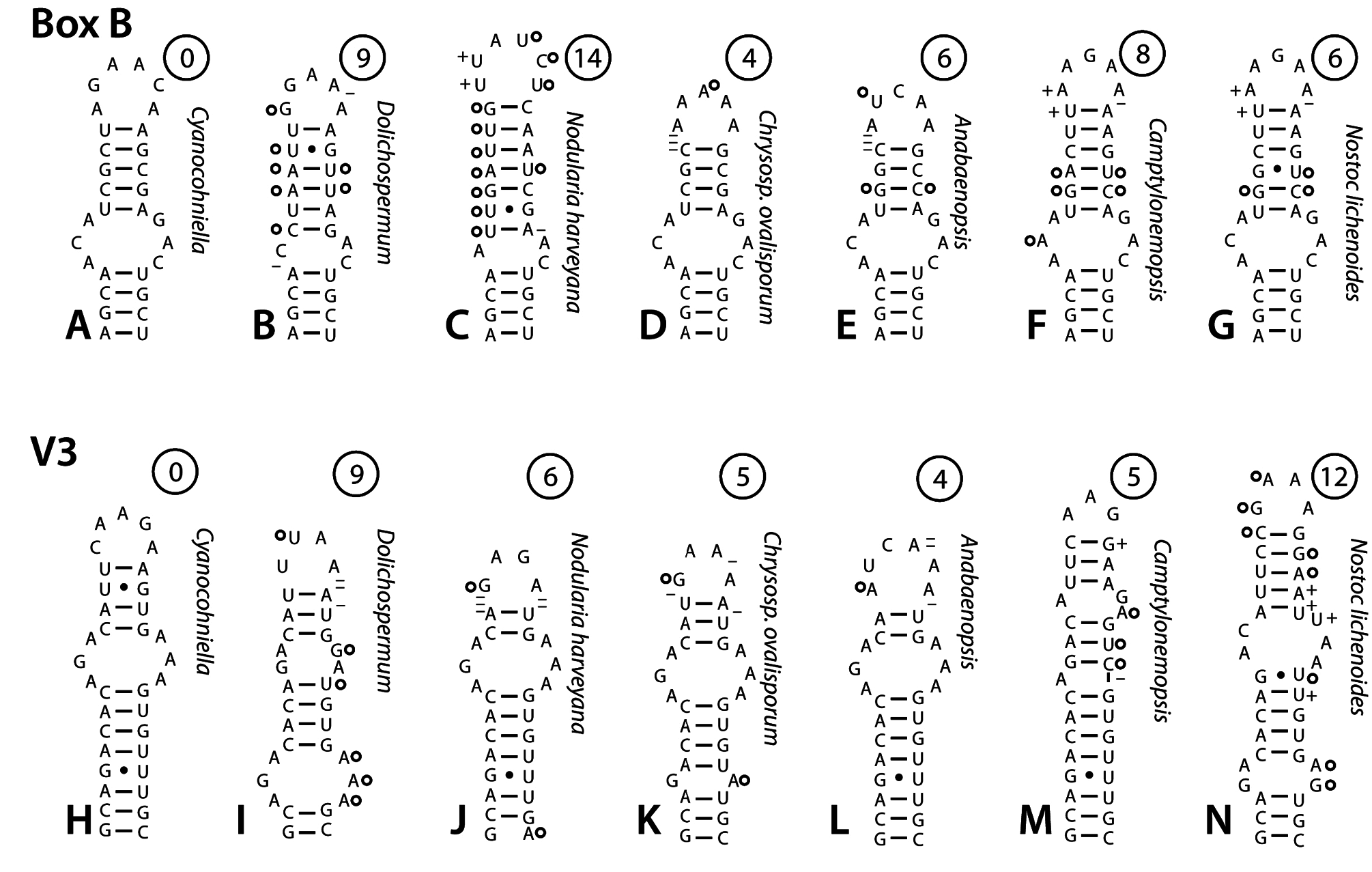
Secondary structure of the 16S–23S ITS: —Comparison of the secondary structure of the 16S–23S ITS conserved domains provides further evidence of genetic separation from other genera in the Aphanizomenonaceae . The D1–D1’ helix of Cyanocohniella is diffrent from all D1–D1’ helices in related genera in the Nostocales ( Fig. 4 View FIGURE 4 ). The closest relatives (based on 16S rRNA gene phylogeny) for which ITS sequence is available are Dolichospermum (Ralfs ex Bornet & Flahault 1888: 228) Wacklin, Hoffmann & Komárek (2009: 60) , Nodularia , Chrysosporum , and Anabaenopsis ( Figs. 4B–E View FIGURE 4 ). These structures are notably different, especially in the basal part of the helix which has two bases (5’-AU-3’) on the 5’ strand opposite the basal unilateral bulge in contrast to the single cytosine residue in this position in Cyanocohniella ( Figs. 4A–E View FIGURE 4 ). The apices of the members of Aphanizonenonaceae are variable, but all are different from the apical region of Cyanocohniella . The central portion of the helix has fairly high sequence similarity in all these strains, but differs in structure in notable ways ( Figs. 4A–E View FIGURE 4 ). Camptylonemopsis Desikachary (1948: 46) , which is much more distant based on 16S rRNA gene phylgeny, actually had the D1–D1’ helix most similar to Cyanocohniella ( Fig. 4F View FIGURE 4 ). Nostoc lichenoides Vaucher ex Řeháková, Johansen, Casamatta, Xuesong & Vincent (2007: 484) is as similar in sequence and structure as most of the Aphanizomenonaceae ( Fig. 4G View FIGURE 4 ). The most conserved regions in terms of sequence and structure across all taxa compared was the basal stem of 6 bp and the subterminal adenine-rich bilateral bulge ( Fig. 4 View FIGURE 4 ). For ease in comparison of sequence differences with Cyanocohniella , minimum numbers of nucleotide substitutions and insertions required to obtain the same sequence as Cyanocohniella are given in circles above each helix ( Fig. 4 View FIGURE 4 ). The position of substitutions and insertions shown are hypothetical, and represent the most parsimonius solution. The actual course of sequence change likely required more steps in order to maintain structure in intermediate stages. Furthermore, identical bases in the ITS could be homoplasies, which would also give an impression of fewer steps than actually occurred. Homoplasies have beeen documented in the ITS regions of green algae ( Caisová et al. 2011) and very likely occur in cyanobacteria as well. The Box B helices available were also all notably different from the Box B helix of Cyanocohniella , the only species to have 5 bp in the upper helix ( Fig. 5 A–F View FIGURE 5 ). Chrysosporum ovalisporum ( Forti 1911: 3) Zapomělová et al. ex Zapomělová et al. (2013: 201) , Anabaenopsis , Nostoc lichenoides and Camptylonemopsis shared identical basal secondary structure with Cyanocohniella , and showed the least separation in number of nucleotide substitutions, insertions, and deletions required to match the sequence of Cyanocohniella ( Figs. 5D–G View FIGURE 5 ). Nodularia harveyana was markedly different in sequence and structure ( Fig. 5C View FIGURE 5 ). The V3 helices of the Aphanizomenonaceae all shared a mid-helix bilateral bulge of identical sequence flanked on either side by highly similar nucleotides, but its position in the helix varied ( Figs. 5H–N View FIGURE 5 ). Anabaenopsis , Chrysosporum ovalisporum , Camptylonemopsis , and Nodularia harveyana had minimal sequence divergence from Cyanocohniella ( Figs. 5J–M View FIGURE 5 ). Nostoc lichenoides , which is typical of the genus Nostoc , was very different in sequence in the V3 helix. The V2 region (not shown) was only available for four strains, as Nodularia and Anabaenopsis lacked tRNA genes, and C. ovalisporum lacks the V2 even though both tRNA genes are present. When all ITS conserved domains are taken into consideration, Cyanocohniella is most similar among the Aphanizomenonaceae strain structures to Chrysosporum ovalisporum and Anabaenopsis . However, Camptylonemopsis which clearly places in the Nostocaceae based on 16S rRNA gene phylogeny, was also quite similar. Cyanocohniella is unique among all Nostocales for which we have presently examined ITS structures.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |