
Coptotriche parvisacculata Diškus & Stonis, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4691.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:14CC7B3E-ACBB-4770-A9D2-3AD35A1A2532 |
|
DOI |
https://doi.org/10.5281/zenodo.5930149 |
|
persistent identifier |
https://treatment.plazi.org/id/767FC3CD-6D2E-4E2D-954A-4132A1233CBE |
|
taxon LSID |
lsid:zoobank.org:act:767FC3CD-6D2E-4E2D-954A-4132A1233CBE |
|
treatment provided by |
Plazi |
|
scientific name |
Coptotriche parvisacculata Diškus & Stonis |
| status |
sp. nov. |
Coptotriche parvisacculata Diškus & Stonis View in CoL , sp. nov.
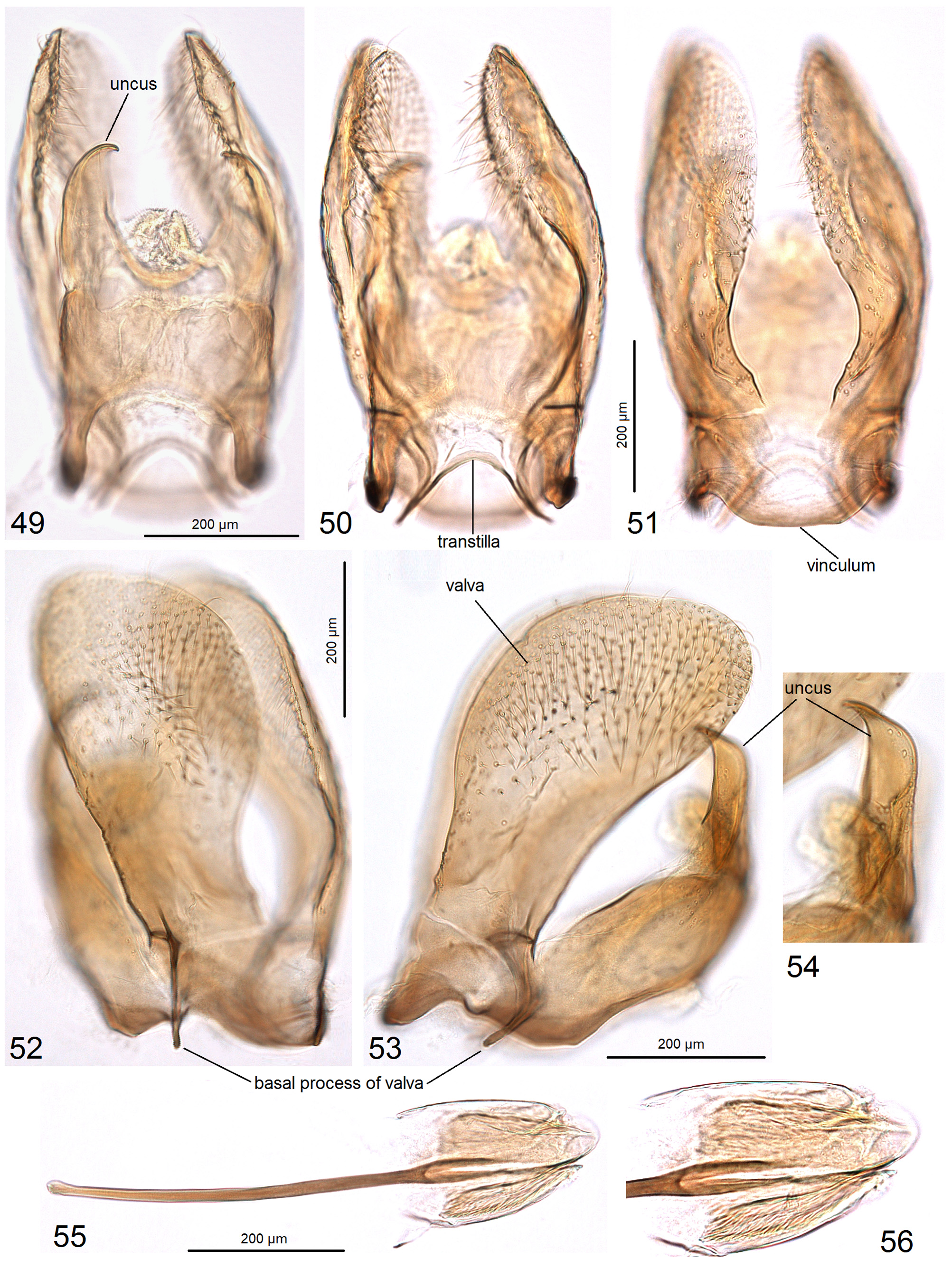
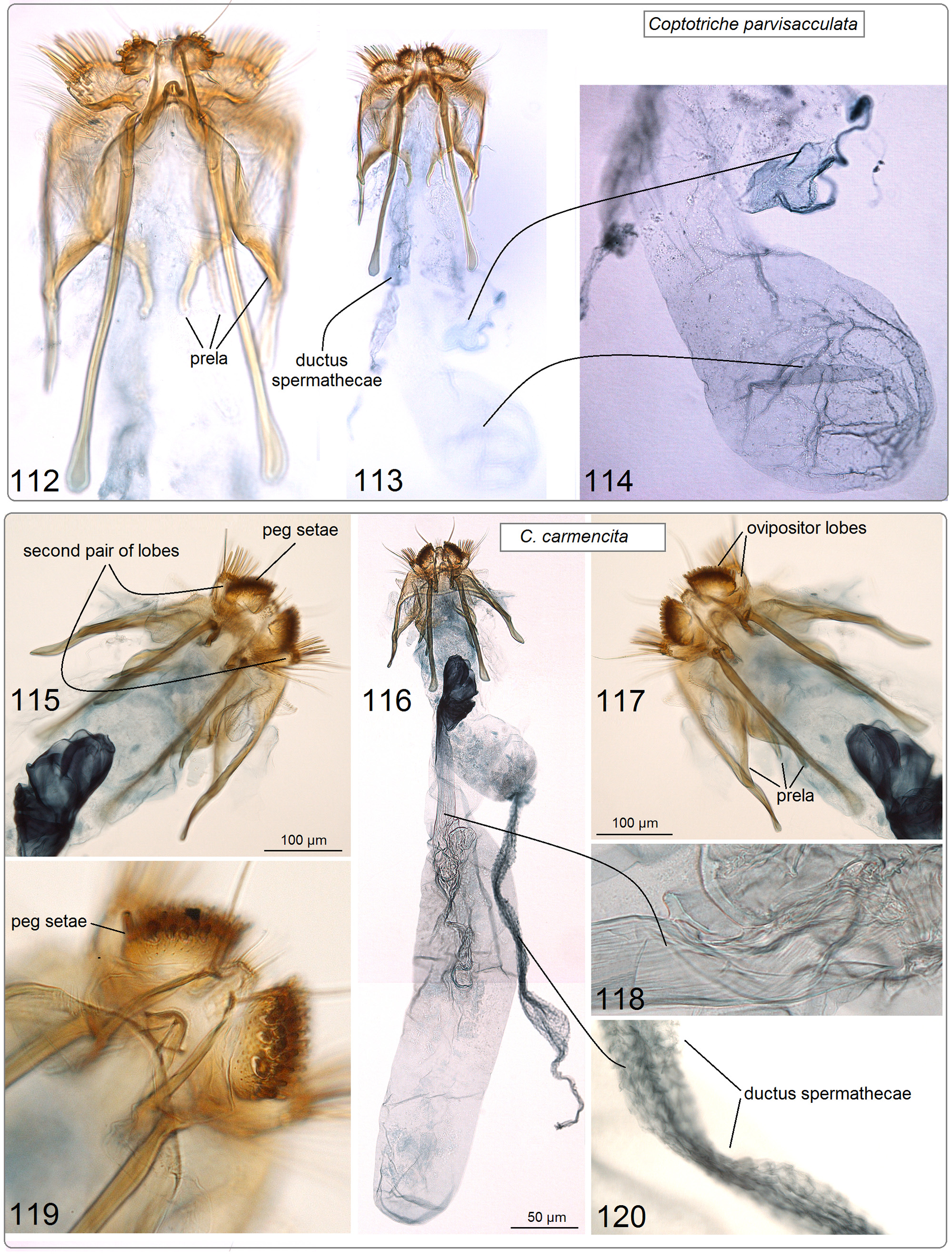
( Figs 1–5 View FIGURES 1–5 , 22 View FIGURES 19–24 , 49–56 View FIGURES 49–56 , 112–114 View FIGURES 112–120 )
Type material. Holotype: ♀, ARGENTINA : Buenos Aires, 34 ° 35ꞌ59ꞌꞌS, 58 ° 21ꞌ3ꞌꞌW, elevation 0 m, mining larvae on Terminalia australis Cambess. ( Combretaceae) , 2.iii.2019, ex pupa iii.2019, field card no. 5305, A. Diškus, genitalia slide no. AD 965♀ ( ZMUC) . Paratypes: 1 ♂, 1 ♀, same label data as holotype, genitalia slide no. AD 966♂ (from an adult in pupal skin) ( ZMUC).
Diagnosis. External characters are not sufficient for species identification. In the male genitalia, the combination of a distinctly truncated vinculum ( Fig. 51 View FIGURES 49–56 ) and a uniquely shaped phallus with weakly developed spines ( Fig. 55 View FIGURES 49–56 ) distinguishes C. parvisacculata from other Coptotriche species, including the South Asiatic C. terminaliae Puplesis & Diškus. In the female genitalia, the elaborate prela ( Fig. 112 View FIGURES 112–120 ) and bulbous corpus bursae ( Fig. 114 View FIGURES 112–120 ) appear to be unique to this species. These female characters may not remain valid for species differentiation because females of many other Coptotriche species are unknown. The host plant, Terminalia australis , and its distinctive leaf mines ( Figs 3–5 View FIGURES 1–5 ) make this species unique in the Neotropical fauna.
Description. Male. Known only from an adult in a pupal skin; for external description see female below. Genitalia ( Figs 49–56 View FIGURES 49–56 ) with capsule about 455 µm long. Uncus with two very large lateral lobes ( Figs 49, 53, 54 View FIGURES 49–56 ). Valva wide, undivided ( Figs 52, 53 View FIGURES 49–56 ), about 505 µm long (excluding the basal process); transtilla short, with distinctive sublateral processes ( Fig. 50 View FIGURES 49–56 ); basal process of valva short ( Figs 52, 53 View FIGURES 49–56 ). Anellus membranous, indistinctive. Diaphragma little spined. Vinculum truncated ( Fig. 51 View FIGURES 49–56 ). Phallus about 530 µm long, strongly broadened in distal one-fourth, with weakly developed spines ( Figs 55, 56 View FIGURES 49–56 ).
Female ( Fig. 22 View FIGURES 19–24 ). Forewing length 3.2–3.3 mm; wingspan 7.3–7.4 mm (n = 2).
Head. Face, labial palpus and pecten yellowish cream; frontal tuft and collar pale grey proximally, yellowish cream distally; antenna distinctly longer than one-half length of forewing; flagellum pale grey to grey cream, at base ochre cream.
Thorax. Tegula, thorax, and forewing concolorous, glossy ochre, with some black scales along forewing margins and at apex; black scales without purple iridescence; fringe grey, with incomplete and inconspicuous fringe line; forewing underside covered with dark grey scales, except forewing apex ochre cream; no androconia. Hindwing pale grey on upper side and underside, without androconia; fringe pale grey. Legs metallic grey, with some yellowish cream scales and bristles.
Abdomen. Dark grey on upper side, grey proximally, yellowish cream distally on underside; no anal tufts. Genitalia ( Figs 112–114 View FIGURES 112–120 ) about 1780 µm long. Ovipositor lobes large with short, stout and darker, modified setae (‘peg setae’) ( Fig. 112 View FIGURES 112–120 ); area between ovipositor lobes indistinct, with tiny papillae and some short setae. Second pair of lobes, lateral and anterior to ovipositor lobes, much larger than ovipositor lobes, bearing very long slender setae, without peg setae ( Fig. 113 View FIGURES 112–120 ). Posterior apophyses much longer than anterior ones ( Figs 112, 113 View FIGURES 112–120 ); prela comprised of three pairs of projections ( Figs 112, 113 View FIGURES 112–120 ). Corpus bursae bulbous ( Fig. 114 View FIGURES 112–120 ), without pectinations or signum. Accessory sac wide; ductus spermathaecae large, without coils.
Bionomics ( Figs 1–5 View FIGURES 1–5 ). Host plant is Terminalia australis Cambess. (Combretaceae) ( Figs 1, 2 View FIGURES 1–5 ). Larvae mine leaves in early March; based on pupae and vacant leaf mines, mining may start as early as February; voltinism unknown. The larva is pale green with a pale brown head and dark green intestine. The blotch mine ( Figs 3–5 View FIGURES 1–5 ) is irregular, usually pale, but with little irregularly deposited frass; fully developed mines may bend (distort) the mined leaf ( Fig. 5 View FIGURES 1–5 ). Adults occur in March.
Distribution. The species is known from the single locality, Buenos Aires, Argentina , but the host plant has a much broader distribution (see Discussion).
Etymology. The species name is derived from the Latin parvus (short or small) and sacculus (a distal part of vinculum) referring to the unique, small and truncated vinculum in the male genitalia.


| ZMUC |
Zoological Museum, University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |