
Copelatus martini, Manuel & Deler-Hernández & Megna & Hájek, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4399.3.7 |
|
publication LSID |
lsid:zoobank.org:pub:3A3D7EB9-3C4B-44EA-8CA8-037361101204 |
|
DOI |
https://doi.org/10.5281/zenodo.6488905 |
|
persistent identifier |
https://treatment.plazi.org/id/5B015862-4932-FF91-D3C6-FB788CCFDDF7 |
|
treatment provided by |
Plazi |
|
scientific name |
Copelatus martini |
| status |
sp. nov. |
Copelatus martini View in CoL sp. nov.
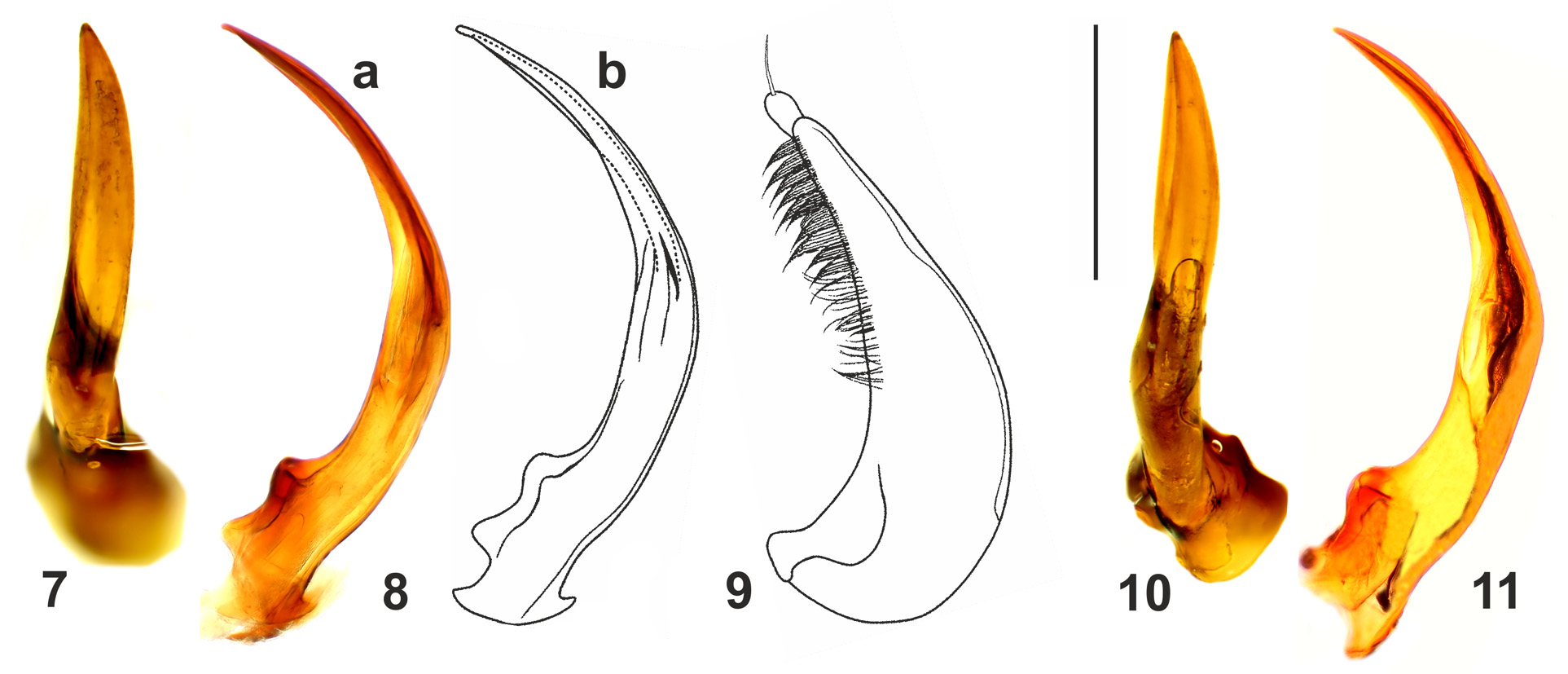
( Figs 1–5 View FIGURES 1–6 , 7–9 View FIGURES 7–11 )
Type locality. Hispaniola Island, Dominican Republic, La Vega, Reserva Científica Ebano Verde , 19°1.96'N 70°32.59'W, altitude 1,070 m a.s.l. GoogleMaps
Type material. Holotype ♂ (NMPC), labelled: "DOMINICAN REP.: La Vega / Res. Ebano Verde (around admin. / centre); 19°1.96'N 70°32.59'W; / 1070 m; 24.viii.2014 / Deler, Fikáček, Gimmel DR20" [printed] // "HOLOTYPE / COPELATUS / martini sp. nov. / det. Manuel et al. 2017" [red, printed]. Paratypes (in total 9♂, 8♀) (CZCTR, MHND, MMCP, NMPC, ZSMG): 6♂, 7♀ (including DNA vouchers MF1021.1 and MF1021.2), same data as holotype; 3♂, 1♀ (including DNA voucher MF1020): "DOMINICAN REP.: Barahona / MN Domingo Fuerte "Cachote" / 18°5.91'N 71°11.35W; 1188 m / 14.viii.2014 / Deler, Fikáček, Gimmel DR03". All paratypes with a respective yellow label.
Description of male holotype. Habitus oblong-elongate; lateral sides subparallel, posteriorly rather strongly attenuated to narrowly rounded apex; lateral outline continuous between pronotum and elytra; maximum body width at about half of total length; dorsal surface weakly convex, medially in anterior third of elytra almost flat.
Colouration. Dorsal surface of head rufo-testaceous, on clypeus vaguely paler, behind eyes darker. Pronotum dark rufous to brown; with diffusely delimited testaceous marginal area, wider near anterior angle, posteriorly narrowing, not reaching posterior angle. Elytra pale brown; along basal margin with vague and narrow paler band, transversally extending from lateral margin to shortly before suture; also vaguely paler along lateral margin in anterior third. Venter of head, pronotal epipleura and median region of prosternum testaceous; rest of ventral surface rufo-testaceous, except brownish abdominal ventrites IV–VI. Appendages testaceous.
Head. Anterior outline of clypeus broadly truncated, slightly concave in middle; on each side of truncation smoothly angulated then obliquely straight until slight curvature in front of eye. Eyes small (ratio between maximum transverse width of left eye and interocular distance about 0.21). Surface strongly shiny, with distinct isodiametric reticulation; with fine, shallow and rather sparse punctures. Sublaterally with two short transverse rows of coarse setigerous punctures in shallow but distinct depressions, first on frons at level of eye anterior margin, second on fronto-clypeal border. Along inner and anterior parts of eye margin with rather deep narrow semi-circular depression containing small coarse setigerous punctures.
Pronotum. Anterior margin weakly arcuate. Posterior margin almost straight. Lateral margins moderately convergent, in posterior half weakly arcuate, in anterior half almost straight; with narrow bead, distinct from posterior angle to level of anterior margin, then vanishing. Maximum width shortly before posterior angles. Surface strongly shiny; with isodiametric reticulation, more weakly impressed than on head; punctation as on head. Laterally on disc, some punctures a bit deeper, in part confluent, forming short, shallow and irregular longitudinaloblique scratches. Along anterior margin with transverse irregular row of coarse and rather dense setigerous punctures, shortly interrupted in middle. Sublaterally along posterior margin, with few coarse setigerous punctures. With well-defined depression along lateral margins, containing small coarse setigerous punctures; depression starting shortly behind anterior angle, longitudinally orientated until posterior third of pronotum length, then deviating obliquely inwards, ending shortly before posterior margin.
Elytron with 10 dorsal striae. Striae 1–7 and 9–10 well impressed and complete or almost complete; stria 8 reduced to short fragments in anterior half of elytron, missing in posterior half; striae 3, 5 and 7 distinctly more impressed than remaining striae; striae 2 and 10 on both elytra and stria 6 on left elytron shortly interrupted close to anterior extremity. Striae 3–8 and 10 starting shortly behind basal margin; striae 1–2 starting slightly more posteriorly; striae 9 starting even more posteriorly. Striae 1, 3, 5, 7, 9 ending shortly before apex; striae 2, 4, 6 and 10 ending at about third quarter of elytral length. Along dorsal striae with very few indistinct large setigerous punctures. Submarginal stria missing, but irregular groups of coarse setigerous punctures present along lateral margin in posterior two thirds of elytra. Surface of scutellum and of elytron between striae strongly shiny, with weakly impressed isodiametric reticulation; meshes smaller than on pronotum; with rather sparse shallow punctures of irregular size; larger punctures fairly distinct, diameter equalling about 2–3 meshes of reticulation; without setae. Subapically behind posterior extremity of striae with larger coarse punctures each bearing one long seta.
Ventral surface. Entirely very shiny except on gula and mediolateral parts of prosternum. Surface of gula with strongly impressed isodiametric reticulation; genae in part smooth and in part with obsolete reticulation. Anterior margin of prosternum medially strongly produced forwards, bearing tuft of very long golden setae. Prosternum surface medially strongly convex, with obsolete reticulation, without setae; sublaterally microgranulose and with long, rather dense golden setae; laterally with obsolete reticulation, impunctate. Surface of prosternal process entirely smooth, almost impunctate. Blade of prosternal process broadly oval, posteriorly strongly attenuated to narrowly rounded apex; maximal width about twice minimum width (between procoxae); with distinct lateral bead continuous from base to apex; with surface evenly and weakly convex, more strongly so towards apex. Elytral epipleura with obsolete reticulation, almost impunctate. Metaventrite and metanepisternum with obsolete reticulation, meshes isodiametric to slightly transverse; almost impunctate. Surface of metacoxa with weakly impressed reticulation, almost impunctate. Metacoxal plates with meshes of reticulation indistinctly elongate in longitudinal-oblique direction; with rather shallow, short longitudinal-oblique scratches. Metacoxal lines weakly divergent, with anterior extremity separated from posterior margin of metaventrite by distance approximately equivalent to maximum width of prosternal apophysis. Lobes of metacoxal posterior margin rounded, with short narrow postero-internal incision. Surface of abdominal ventrites with very obsolete reticulation, somewhat more distinct on ventrite VI, and with fine to medium-sized shallow punctures; orientation of meshes longitudinal on ventrite I–II, oblique on ventrites III–IV, transverse on ventrite V; isodiametric on ventrite VI. At midline of ventrites III–V and sublaterally on both sides of ventrite VI with isolated group of few coarse punctures bearing long golden setae.
Appendages. Antennae rather long; antennomeres II–X regularly broadened from proximal to rather thick distal extremity, slightly more than twice as long as broad. Anterior margin of protibia straight, not emarginated. Pro- and mesotarsi rather broadly dilated, on ventral surface with 4 transverse rows of 3 (most proximal row) or 4 (other rows) of oval suction setae. Inner and outer claws of pro- and mesotarsi equal, rather thick; outer claw more strongly curved in proximal half than in distal half; inner claw more evenly curved.
Aedeagus. Median lobe in lateral view sickle-shaped with bluntly pointed apex ( Fig. 8 View FIGURES 7–11 ); in ventral view as in Fig. 7 View FIGURES 7–11 ; paramere as in Fig. 9 View FIGURES 7–11 .
Females. Pro- and mesotarsi not broadened; pro- and mesotarsal claws narrower than in males. Dorsal sculpture dimorphic; homeomorph form with dorsal sculpture as in male; heteromorph form with strongly developed sexual structure: most of head surface with short strioles, transverse in anterior half of head and longitudinal in posterior half; pronotum surface entirely and densely covered with deep long strioles, elongated mainly in longitudinal direction, somewhat sinuate, partly anastomosing; elytral surface except in posterior quarter entirely covered by long longitudinal anastomosing strioles, these more densely and regularly spaced than on pronotum; elytral surface more matt than on head and pronotum, with reticulation more strongly impressed; elytral striae distinct but very narrow and superficial.
Variability. In addition to the dimorphism of female dorsal sculpture described above, there is noticeable variation, in both males and females, at the intra- and inter-population levels. Within populations, specimens vary slightly in colour pattern, the lateral pale area on pronotum and the basal band on elytra being more or less developed, in some specimens with the latter at least twice broader close to the scutellum than laterally. Strong variation affects the sculpture of the pronotal surface in the lateral regions of the disc (in males and in homeomorph females). Some specimens display limited confluence of punctures and, therefore, only few shallow rugae (as in the holotype), whereas others have in addition a few "true" short strioles (one male paratype and one female paratype), or even display rather dense short strioles throughout pronotum surface except in the central region of the disc (the other female paratype).
Furthermore, we observed remarkable morphological variation between the two different populations of this species. In the " La Vega " population (to which the holotype belongs), specimens have 10 dorsal elytral striae (but with stria 8 strongly abbreviated posteriorly, in most specimens fragmented, or even reduced to a few residual scratches), and the basal testaceous band of elytra is comparably narrow. In the "Cachote" population, the even dorsal striae are much more reduced. Two of the male paratypes from this locality have only sparse remnants of striae 2, 4, 6, 8. In the other male paratype and in the single female, striae 2, 4, 6, 8 are totally lacking, and, therefore, these specimens have only 6 dorsal striae (equivalent to striae 1, 3, 5, 7, 9 and 10 of the other specimens). In addition, all four paratypes from "Cachote" have the basal band of elytra broader and of a brighter testaceous colour. Of note is that the single female paratype collected from the "Cachote" population has heteromorph dorsal sculpture, but both female morphs are represented among the " La Vega " specimens.
Measurements. Holotype: total length (TL) = 4.7 mm, total length without head = 4.2 mm, maximum width (MW) = 2.1 mm, ratio TL/MW = 2.24. Paratypes: TL = 4.6–5.1 mm (4.85 ± 0.19), TL without head = 4.0– 4.6 mm (4.30 ± 0.20), MW = 2.05–2.45 mm (2.26 ± 0.13), TL/MW = 2.08–2.24 (2.15 ± 0.06).
Differential diagnosis. Copelatus martini sp. nov. belongs to the consors -group of Guignot (1961) and Guéorguiev (1968) (= group 5 of Sharp 1882), defined by the absence of a submarginal stria and the general presence of 9 or 10 dorsal striae (we consider the presence of only six elytral striae in two specimens from "Cachote" as individual variability). Including the new species, the consors -group currently comprises 20 species, including 13 Neotropical ones. Indeed, to the 18 species traditionally listed in must be added not only C. martini sp. nov., but also C. guadelupensis Legros, 1948 —a species erroneously listed in the erichsonii -group by Guéorguiev (1968), see also Nilsson & Hájek (2018). According to morphological characters Copelatus martini sp. nov. seems to be closely allied to C. guadelupensis . In addition to the diagnostic characters of the consors -group, these two species are similar in habitus and dorsal colour pattern, sculpture of the pronotum surface, reduction of the even dorsal striae, particularly the stria 8 (posteriorly strongly abbreviated, often fragmented), absence of emargination of the antero-basal margin of protibia in male, shape of the median lobe and parameres and existence of two different female morphs. We provide in Table 1 a list of diagnostic characters to help separating these two species, as well as habitus and median lobe illustrations of C. guadelupensis for comparison ( Figs 6 View FIGURES 1–6 , 10, 11 View FIGURES 7–11 ).
These two species are apparently close to the continental Neotropical species C. incognitus Sharp, 1882 (type locality: Mexico, Veracruz, Córdoba) (= neglectus Sharp, 1882; type locality: Venezuela, Sucre state, Cumaná) and C. consors Sharp, 1882 (type locality: Argentina, Pampas). In his description of C. guadelupensis, Legros (1948) mentioned the similarity of that species to C. incognitus ; on the other hand, Sharp (1882: 575) stated that C. consors and C. incognitus are closely allied. According to Sharp's descriptions, the dorsal colour pattern of C. incognitus and C. consors is similar to that of C. martini sp. nov. and C. guadelupensis . The habitus of C. incognitus is furthermore similarly narrow-elongated ( C. consors has a broader body). Copelatus consors is characterised by a reduction of the stria 6 and especially the stria 8, both being in some individuals nearly entirely absent ( Sharp 1882: 575), which is reminiscent of the situation observed in C. martini sp. nov. Finally, both C. incognitus and C. consors have two female morphs, one with the dorsal sculpture as in males, and the other one with the surface of pronotum and elytra covered by very dense strioles.
Analysis of partial CO1 sequences. The molecular data confirms that C. martini sp. nov. is a species distinct from C. guadelupensis ( Fig. 12 View FIGURE 12 ). Uncorrected genetic distances between the sequenced CO1 haplotypes of C. martini sp. nov. and C. guadelupensis are 10.3%–12.5%, well above the highest levels of intraspecific divergence known in dytiscids for this molecular marker (see for example, Bergsten et al. 2012). In the CO1 tree, C. martini sp. nov. and C. guadelupensis are not even sister species. Several undetermined Copelatus species (" Copelatus sp." in Fig. 12 View FIGURE 12 ) sequenced in previous studies are closer to C. guadelupensis than is C. martini sp. nov. In the CO1 tree, C. distinctus Aubé, 1838 is sister to C. martini sp. nov. (without statistical support). Haplotypes of C. guadelupensis from Guadeloupe and from Puerto Rico fall close to each other but are relatively distant (4.1%– 4.7%). Furthemore, those from Guadeloupe form an unsupported clade with two undetermined sequences from Brazil and Argentina respectively. Morphological examination by two of the co-authors (YM and JH) of the DNA vouchers of these specimens (MB0620 and MB7035, deposited in ZSMG) led to the conclusion that these specimens do not belong to C. guadelupensis . We consider for the moment, that based on morphology specimens from Guadeloupe and Puerto Rico belong to the same species ( C. guadelupensis ), which is therefore paraphyletic in the CO1 tree (not an uncommon situation, see Bergsten et al. 2012). However, this clearly requires more indepth study through sequencing of more specimens and/or analyses of additional molecular markers.
C. martini sp. nov. C. guadelupensis
Size on average slightly smaller; 4.6–5.1 mm on average slightly larger; 4.8–5.5 mm
Colour of ventral surface mainly rufo-testaceous slightly darker (mainly ferrugineous)
Anterior outline of clypeus on each side of broad medial truncation on each side of medial truncation more evenly
somewhat angulated, posteriorly straight rounded
Fronto-clypeal depressions well impressed indistinct
Eye size smaller (ratio between maximum transverse slightly larger (ratio between maximum width of left eye and interocular distance transverse width of left eye and interocular about 0.21–0.23) distance about 0.24–0.25)
Longitudinal depression along well impressed very shallow, almost indistinct
pronotum lateral margins
Elytral striae stria 8 reduced, fragmentary or missing; stria 8 reduced, but to lesser extent; striae 2, 4,
striae 2, 4, 6 in some specimens fragmentary 6 present and complete
or missing
Surface of blade of prosternal evenly and rather weakly convex more strongly convex; almost tectiform in
process cross-section
Metacoxal lines anteriorly ending well before posterior longer; ending shortly behind posterior margin of metaventrite margin of metaventrite
Aspect of dorsal surface strongly shining; reticulation on surface of slightly less strongly shining; reticulation on pronotum and elytra weakly impressed; surface of pronotum and elytra slightly more posterior part of elytra almost without impressed; on posterior part of elytra with iridescent sheen strong iridescence (blue-green sheen)
Median lobe in lateral view ventral outline evenly curved; outline near ventral outline less evenly and slightly less basal piece and near apex forming a right strongly curved; angle between ventral outline angle; subapically somewhat narrowed then near basal piece and near apex more than 90°; slightly dilated before narrowly rounded apex distally evenly narrowed to pointed apex
Median lobe in ventral view rather thick; left outline weakly sinuous; narrower; left outline more strongly sinuous; apical region weakly asymmetric apical region more strongly asymmetric (bent left)
Etymology. The specific epithet martini is a noun in the genitive singular, chosen in honour of our excellent friend Martin Fikáček (National Museum in Prague) for his contribution to the study of aquatic insects. Collection circumstances. Copelatus martini sp. nov. was collected in temporary lagoons with turbid water, with and without aquatic vegetation, with abundant vegetal detritus and muddy bottom in the mountain zone (1,070 and 1,188 m respectively).
Distribution. Copelatus martini sp. nov. is thus far known only from the Dominican Republic, in the southern (Paleoisla Sur) and central parts (Paleoisla Norte) of the country ( Fig. 13 View FIGURE 13 ).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Copelatinae |
|
Genus |