
Ceropegia gilboaensis Heiduk, N.R.Crouch & D.Styles, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.591.2.4 |
|
DOI |
https://doi.org/10.5281/zenodo.7871074 |
|
persistent identifier |
https://treatment.plazi.org/id/03B2821F-FF96-FFD8-7782-9092031EFA3F |
|
treatment provided by |
Ronellklopper |
|
scientific name |
Ceropegia gilboaensis Heiduk, N.R.Crouch & D.Styles |
| status |
sp. nov. |
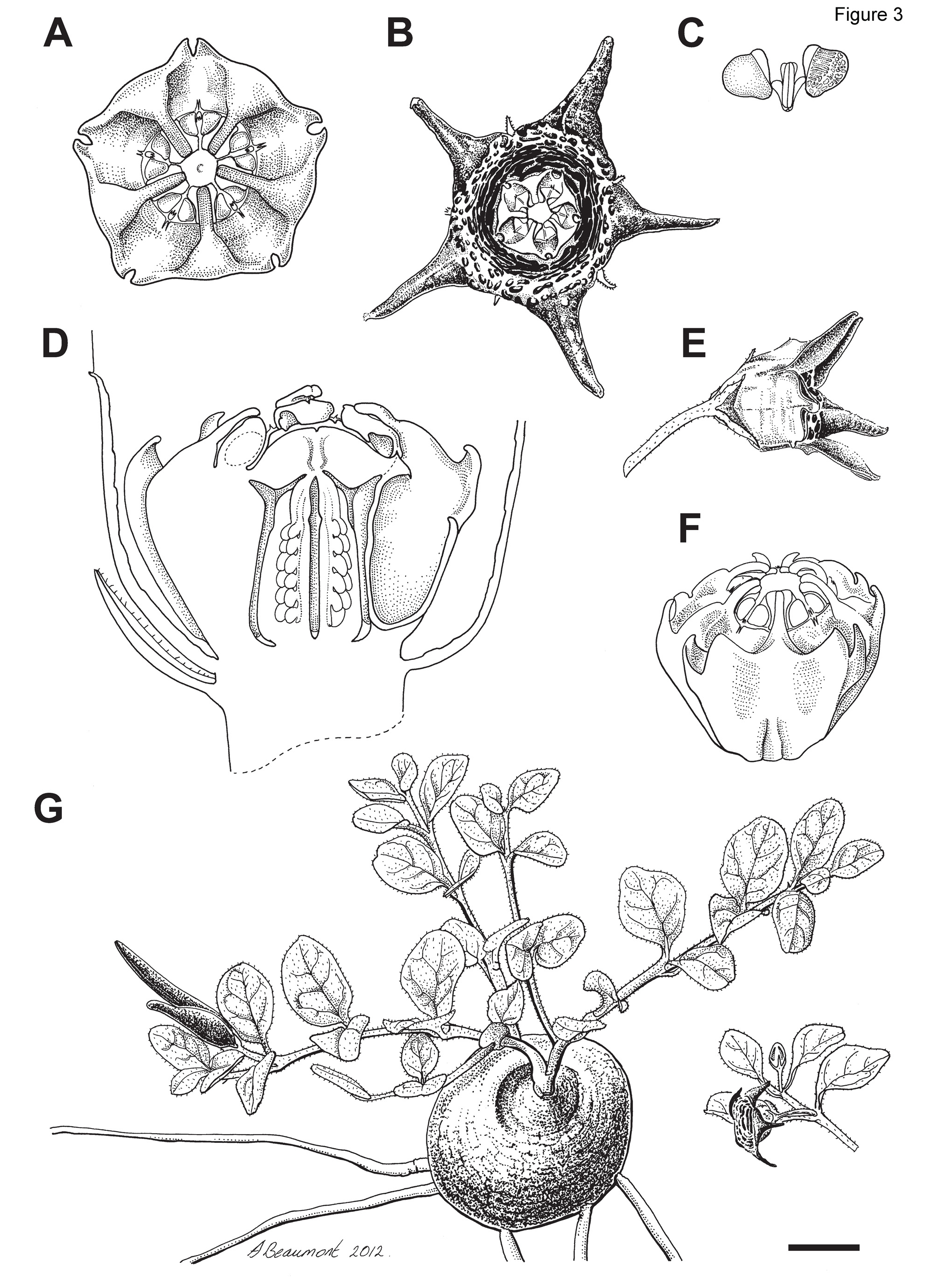
Ceropegia gilboaensis Heiduk, N.R.Crouch & D.Styles View in CoL sp. nov. ( Figs 3 View FIGURE 3 , 4 View FIGURE 4 A−E, 5A)
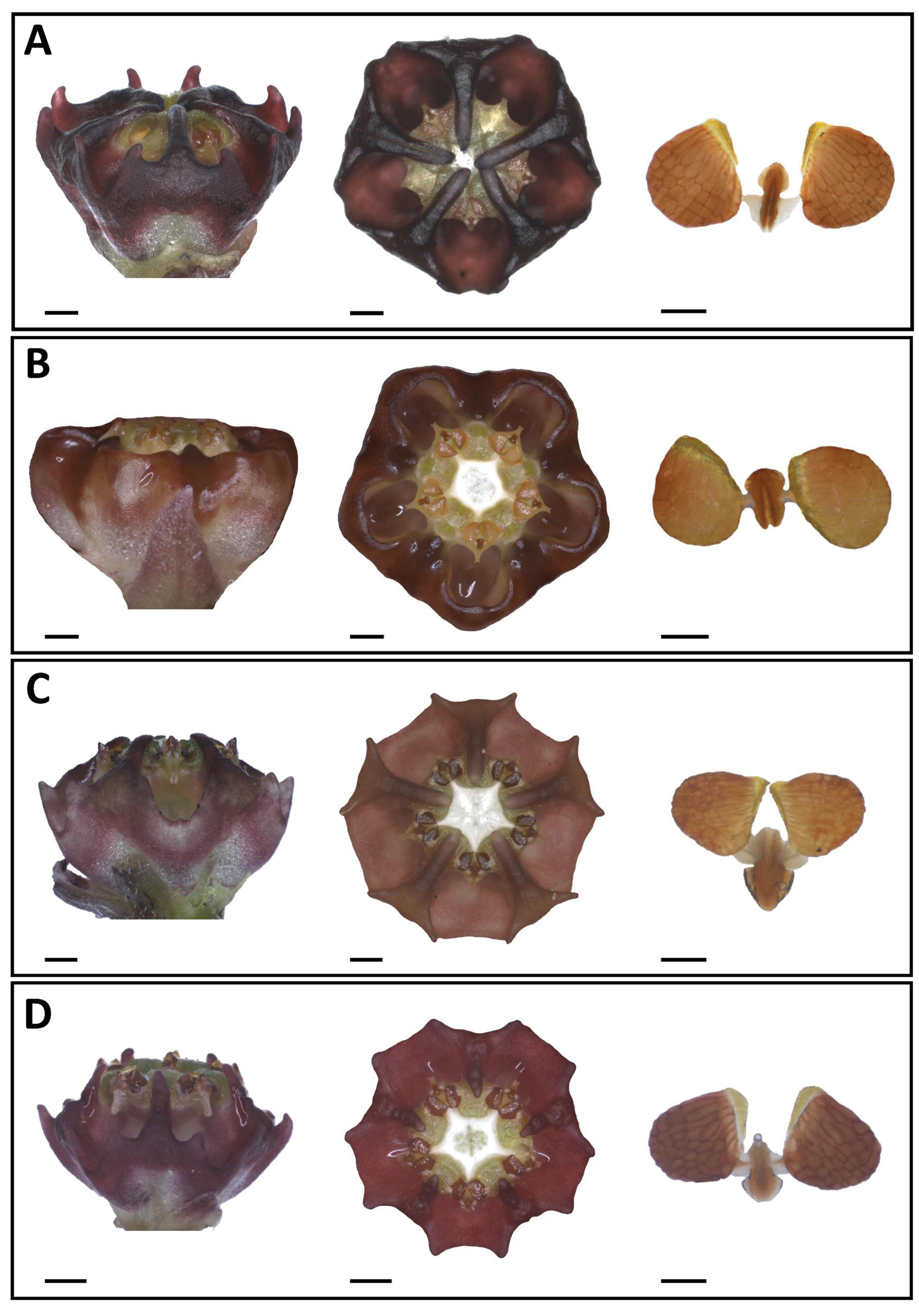
Diagnosis: —Plants of Ceropegia gilboaensis are perennial tuberous succulents with a prostrate growth habit, ovate to elliptic leaves, verrucose follicles, and a cylindrical-campanulate corolla, that can be separated from C. ngomensis ( Figs 1I View FIGURE 1 , 5B View FIGURE 5 ) in lacking the distinctly keeled or fluted corolla lobe bases that embed the gynostegium forming a characteristic 5-star shaped corolla mouth in C. ngomensis . The basally fused corolla lobes of C. gilboaensis have transverse burgundy bands but are purple-mottled in C. ngomensis . In C. gilboaensis , the inner corona lobes are adpressed to the back of the stamens along their full length and closely proximate to subconnivent over the style-head, whilst in C. ngomensis they are short and only adpressed to the base of the stamens. The outer corona lobes are deeply bifid in C. gilboaensis but undivided to form five cups with incurved margins in C. ngomensis .
Type: — SOUTH AFRICA. KwaZulu-Natal, the Karkloof, Mt Gilboa , ca. 1620 m, 12 November 2020, D. G. A. Styles & A. Heiduk 5841 (holotype; NU0092539 !) .
Description: —Perennial dwarf herb. Root a subterranean tuber, 20–40 mm in diameter, ca. 20 mm high, somewhat depressed on upper side, rounded below. Stems 1 or 2, up to 150 mm long, produced annually, branched sparingly at base, slender, creeping, prostrate, shortly pubescent. Leaves in 3–6(–8) pairs, shortly petiolate (to 1.7 mm long), lamina elliptic-ovate, 10–12 × 7–9 mm, ciliate with white hairs along the thin often purplish margins, pubescent above and below, venation prominent on lower surface and recessed on upper surface, basally rounded, apically acute. Flowers 1(–2) lateral at nodes, open in succession, with pungent fetid, slightly acidic-fecal floral scent; pedicels slender, pubescent, 6–7 mm long. Sepals narrowly triangular, 2.2–3.7 mm long, sparsely pubescent. Corolla steep cylindrical-campanulate, ca. 13–14 mm across, corolla lobes spreading, divided into fused and unfused section ± halfway; tube 4.5–6.3 mm deep, velvety, glabrous (rarely with sparse white trichomes on margins of corolla lobe tips), cream within with raised, transverse burgundy bands that proximally are more entire than distally; lobes narrowly triangular, 3.3–5.4 mm long, keeled, concave above, proximally cream with raised burgundy striations, distally velvety and brownish-red to rarely olive-green. Gynostegium sessile, glabrous, dark purple. Gynostegial corona 3.6–4.4 mm in diam., 2.0– 3.1 mm high, of staminal and interstaminal parts, interstaminal (outer) corona lobes joined to form a cup with V-shaped outer margin, deeply bifid, lobules ca. 0.3 mm long, falcate, inclined towards the inner lobes; staminal (inner) corona lobes adpressed to stamina along their entire length and incumbent on style-head, linear, ca. 1.7 × 0.3 mm, apices inclined over the staminal column, closely proximate to subconnivent. Pollinarium pollinia sagittate, ca. 450 × 400 μm, brownish, insertion crest ca. 250 μm long, stramineous; caudicles ca. 90 μm long; corpusculum sagittate, ca. 315 × 140 μm, brown. Follicle with both mericarps developing, erect, ca. 40–45 mm long and 4 mm at widest point, verrucose, green flecked with purple, maturing to purple-brown. Seeds ovate-elliptic, ca. 6 × 2 mm, brown, comose, coma ca. 7 mm, white.
Additional material examined:
Ceropegia gilboaensis : SOUTH AFRICA. KwaZulu-Natal, the Karkloof, Mt Gilboa , ca. 1630 m, 03 April 2022, D. G. A. Styles & A. Heiduk 6032 ( NU0092540 !; fruiting material) .
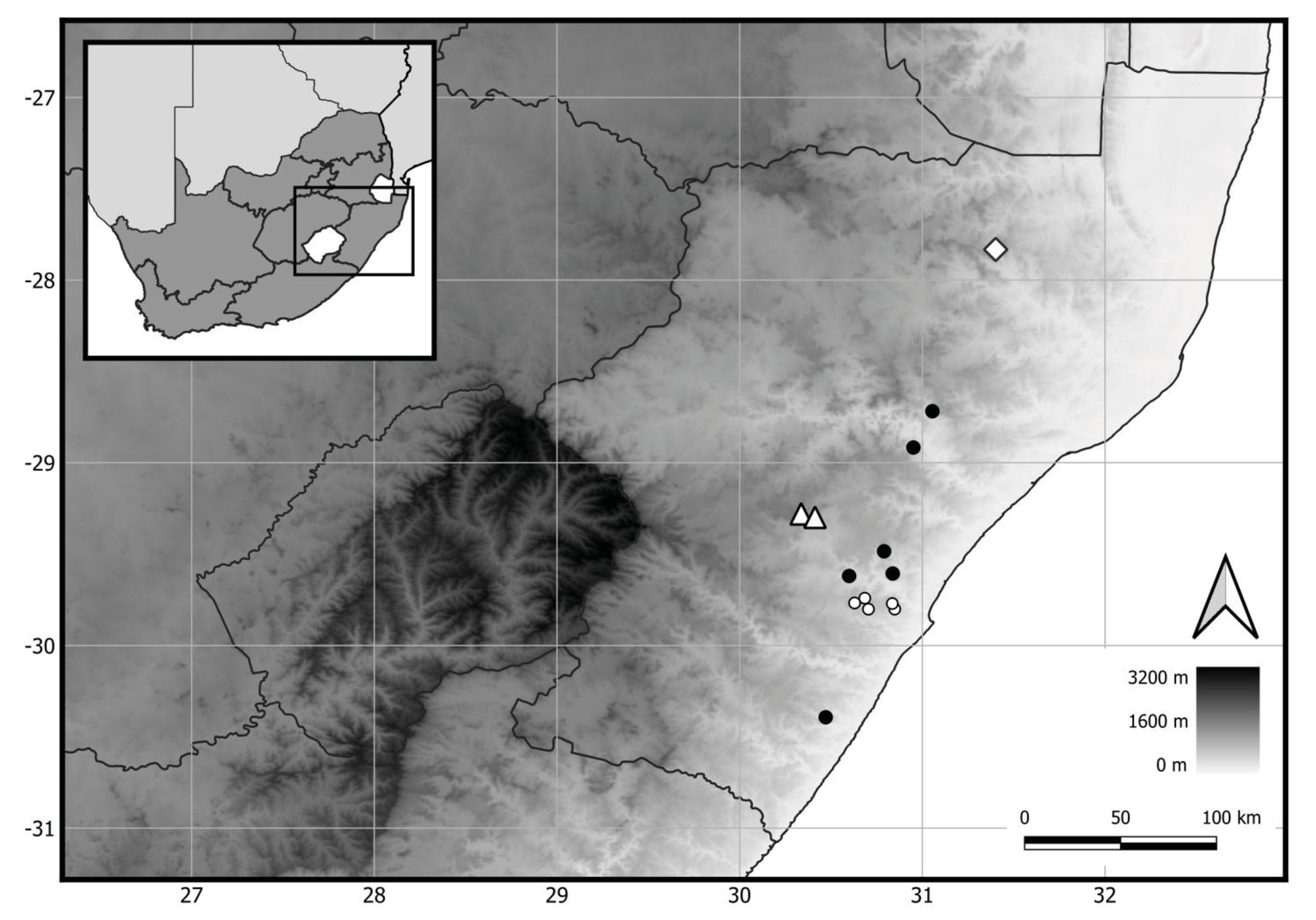
Distribution and ecology: — Ceropegia gilboaensis is to date known only from the Karkloof in the KwaZulu-Natal Midlands, South Africa. It is the only representative of the genus documented from the Karkloof, and is considered endemic there, at elevations of 1320−1670 m. This new species is found within grassland, growing exposed in shallow soil on the margins of, and in between, dolerite rock outcrops. Common grasses at the sites include Aristida junciformis Trin. et Rupr. (1842: 143) and Sporobolus natalensis ( Steudel 1855: 154) Durand & Schinz (1894: 822) . Associated herbaceous species include Aloe neilcrouchii Klopper & Smith (2010: 95) , Crassula dependens Bolus (1881: 391) and Ledebouria sandersonii ( Baker 1870: App. 5) Venter & Edwards (2003: 50).
The national vegetation map shows that C. gilboaensis occurs in Midlands Mistbelt Grassland and Mooi River Highland Grassland ( South African National Biodiversity Institute 2006–2018). In this area, Midlands Mistbelt Grassland appears to have been mapped as occurring below a contour approximating an elevation of 1500 m, and Mooi River Highland Grassland above this. However, this likely represents an over-simplification of the floristics in contact areas, where C. gilboaensis occurs just above 1500 m.
Midlands Mistbelt Grassland is described as “forb-rich, tall, sour Themeda triandra [ Forsskal (1775: 178) ] grasslands transformed by the invasion of native Ngongoni grass ( Aristida junciformis subsp. junciformis ). Only a few patches of the original species-rich grasslands remain.” ( Mucina et al. 2006: 422). This vegetation type is “Endangered (one of the most threatened vegetation types of KwaZulu-Natal)” according to Mucina et al. (2006: 423). Furthermore, the Mistbelt of KwaZulu-Natal has been identified as an important centre of endemism ( Mucina et al. 2006), although not yet formally recognized as such ( Mucina et al. 2006; Van Wyk & Smith 2001). More limited floristic information is provided on Mooi River Highland Grassland, which is also considered threatened, but with the status of Vulnerable rather ( Mucina et al. 2006).
Phenology: —Seen in flower from October to December, and in fruit from Jan uary to April.
Etymology: — Ceropegia gilboaensis is named for the type locality at Mt Gilboa in the Karkloof, SouthAfrica, where it was discovered in 2008 by Isabel Johnson, a field botanist, ecological consultant and dedicated conservationist.
Conservation Status: — Ceropegia gilboaensis is only known from two localities 7.5 km apart from each other. At each locality, plants were observed within an area of less than 1 km 2. However, a study of aerial imagery indicates that within the surrounding landscape there may be a further 35 km 2 of similar habitat in which C. gilboaensis could potentially occur. The total number of plants is estimated at less than 250 individuals. The grassland habitat at one of the two localities suffers from the trampling effects of cattle, but does not otherwise appear to be under anthropogenic threat. The species’ conservation status is suggested as Endangered (EN) under IUCN criteria B2(a)(b)(iii) and D ( IUCN Standards and Petitions Subcommittee 2019).
Discussion
Ceropegia gilboaensis is a new species in section Chamaesiphon , the largest section in Ceropegia s.l. with 115 tuberous species ( Bruyns et al. 2017). The overwhelming majority (ca. 90%) of these are species with open non-trapping, brood-site mimicking flowers; species with kettle-trap flowers have evolved in at least three subclades within the section ( Bruyns et al. 2017). Many non-trap flowered species in sect. Chamaesiphon convergently evolved morphological features considered typical for brood-site mimicking plants, i.e., prostrate or sub-erect stems, foul smelling, reddish-purplish or maroon speckled or striated flowers presented close to the ground ( Johnson & Schiestl 2016; Fig. 1A–L View FIGURE 1 ).
Among the brood-site mimicking species in sect. Chamaesiphon , C. gilboaensis appears to be most closely related to C. ngomensis ( Fig. 1I View FIGURE 1 )—a KwaZulu-Natal endemic species only known from the a rea of Ngome, about 220 km north-east of the Karkloof where C. gilboaensis occurs. Ceropegia ngomensis is the only other South African species presenting similarly structured steep cylindrical-campanulate corolla tubes in combination with prostrate stems. Preliminary unpublished molecular findings support a close relationship between both species (Heiduk et al. unpublished). Although C. gilboaensis also shares particular vegetative and floral characters with other brood-site mimicking members of sect. Chamaesiphon occurring in KwaZulu-Natal and adjacent regions, it may be distinguished as explicated below (see also Table 1 View TABLE 1 ).
The prostrate stems with elliptic leaves of C. gilboaensis are distinct from the sub-erect to erect stems with commonly elliptic- or ovate-lanceolate leaves of C. alpina ( Dyer 1977a: 18) Bruyns in Bruyns et al. (2017: 433), C. australis , C. campanuliformis , C. chlorozona , C. modestantha , C. perdita ( Dyer 1977a: 10) Bruyns in Bruyns et al. (2017: 434), C. petrophila and C. spathulata ( Lindley 1827: t. 1113) Bruyns in Bruyns et al. (2017: 435).
The corolla of Ceropegia gilboaensis is, with rare exception (one plant found with sparse white trichomes on corolla lobe margins), glabrous, allowing for separation from C. alpina , C. australis , C. campanuliformis , C. chlorozona , C. perdita , C. petrophila , C. remota , C. swazica ( Dyer 1976: 56) Bruyns in Bruyns et al. (2017: 435) and C. spathulata which all have distinctively puberulous, hirsute or pilose corolla lobes and/or vibratile hairs fringing the corolla lobes ( Fig. 1 View FIGURE 1 , Table 1 View TABLE 1 ; Dyer 1980).
In Ceropegia gilboaensis the corolla is steep cylindrical-campanulate ( Figs 3A–B, F View FIGURE 3 , 4A–D View FIGURE 4 ), which is distinct from the bowl-shaped corolla of C. chlorozona ( Fig. 1F View FIGURE 1 ) and the flat or shallowly saucer-shaped corollas of C. bruceae ( Fig. 1E View FIGURE 1 ), C. coddii ( Fig. 1G View FIGURE 1 ), and C. pulchellior ( Fig. 1K View FIGURE 1 ). Campanulate corollas are also characteristic of C. alpina , C. australis ( Figs 1A–D View FIGURE 1 ), C. modestantha ( Fig. 1H View FIGURE 1 ), C. perdita , C. petrophila ( Fig. 1J View FIGURE 1 ), C. remota ( Fig. 1L View FIGURE 1 ) and C. spathulata . However, in these species the corollas are less steep and the corolla lobes generally proportionally longer than the fused portion of the corolla (ca. ±1/ 2 in C. gilboaensis , see Table 1 View TABLE 1 ); in C. campanuliformis , the corolla lobes are proportionally shorter (±1/3) than the fused part of the corolla.
The follicles of Ceropegia gilboaensis are verrucose, while they are smooth in all other species discussed here. Generally, raucous, non-smooth follicles are rather rare in Ceropegia although known for example from the succulentleaved climbers C. sandersonii Hooker (1869: t. 5792) , which has follicles markedly verrucose to tuberculate, and C. cimiciodora Obermeyer (1933: t. 488) in which species they are thinly tuberculate ( Dyer 1980). Within sect. Chamaesiphon , verrucose follicles have not earlier been reported.
Ceropegia pulchellior and C. modestantha are two KwaZulu-Natal endemics that grow in rocky grasslands in relatively close proximity to C. gilboaensis ( Fig. 2 View FIGURE 2 ), which in view of their superficial similarity could lead to confusion with C. gilboaensis . However, in addition to the differences mentioned above, C. gilboaensis can be distinguished from these species as follows: when viewed from above, the sepals of C. pulchellior protrude from beneath the fused corolla portion, a feature not seen in C. gilboaensis . The spreading corolla lobe tips are basally ovate and distally attenuate-tapering and often reflexed in C. modestantha but are clearly triangular in C. gilboaensis . In C. pulchellior the gynostegium projects from the centre of the corolla whereas in C. gilboaensis , as also in C. modestantha , the gynostegium is sunk into the fused section of the corolla. The margin formed by the outer corona lobes is U-shaped in both C. modestantha ( Fig. 5C View FIGURE 5 ) and C. pulchellior ( Fig. 5D View FIGURE 5 ), while it is V-shaped in C. gilboaensis ( Fig. 5A View FIGURE 5 ). The outer corona lobes are lower than the style-head in C. modestantha ( Fig. 5C View FIGURE 5 ) and C. pulchellior ( Fig. 5D View FIGURE 5 ) but are level with the style-head in C. gilboaensis ( Fig. 5A View FIGURE 5 ).
Both Ceropegia modestantha and C. pulchellior occur on Natal-Group sandstone while C. gilboaensis occurs on dolerite, as does C. ngomensis . Ceropegia modestantha is relatively widely distributed, ranging from near Nkandla in the north, to the Little Noodsberg, and south to the Umzinto coastal escarpment ( Dyer 1980; Styles & Wragg 2008). Ceropegia pulchellior has a more limited distribution than C. modestantha , occurring between Pietermaritzburg and Durban along a narrow line of suitable habitat ( Styles & Wragg 2008), with an outlier subpopulation to the south, “on an isolated mountain top near Eston…” ( Styles & Wragg 2008: 46), within the range of C. modestantha . There, both species apparently co-occur and probably hybridize ( Styles & Wragg 2008). A third species occurring relatively close to C. gilboaensis , but ranging somewhat further to the northeast is C. chlorozona , which grows on dolerite as well as sandstone.
Other species in sect. Chamaesiphon share vegetative and/or floral characteristics with C. gilboaensis but are geographically separated from it. From north to south these species occur as follows: C. bruceae and C. coddii on the Mpumalanga escarpment and in areas of north-eastern eSwatini ( Dyer 1980), with C. coddii also being recorded in KwaZulu-Natal (pers. obs.); C. swazica in eSwatini and adjacent Mpumalanga; C. remota in the Utrecht-Wakkerstroom-Paulpietersburg areas of KwaZulu-Natal; C. petrophila at Boston, Byrne and at Underberg in KwaZulu-Natal; C. perdita in the Drakensberg ( Dyer 1980) above 2000 m near the trijunction of KwaZulu-Natal, Eastern Cape and Lesotho; C. alpina above 2300 m in southern Lesotho; C. campanuliformis and C. spathulata in the Eastern Cape; C. australis at Port Shepstone and as far south as Port St. Johns, restricted to Msikaba Formation sandstone. This lastmentioned species shows remarkable variation in flower colour and the length of the corolla lobes ( Figs 1A–D View FIGURE 1 ; see also Dyer 1983), even within the same population ( Figs 1A–C View FIGURE 1 ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |