
Candobrasilopsis rochai, Higuti & Martens, 2012
|
publication ID |
https://doi.org/ 10.5852/ejt.2012.33 |
|
publication LSID |
lsid:zoobank.org:pub:85998904-3543-461B-A741-CC6959AA48DE |
|
DOI |
https://doi.org/10.5281/zenodo.3859036 |
|
persistent identifier |
https://treatment.plazi.org/id/152E5E0F-0FF9-4A83-B8EA-6424D6A07858 |
|
taxon LSID |
lsid:zoobank.org:act:152E5E0F-0FF9-4A83-B8EA-6424D6A07858 |
|
treatment provided by |
Valdenar |
|
scientific name |
Candobrasilopsis rochai |
| status |
sp. nov. |
Candobrasilopsis rochai sp. nov.
Figs 2-6 View Fig View Fig View Fig View Fig View Fig , 7 View Fig A-B
urn:lsid:zoobank.org:act:152E5E0F-0FF9-4A83-B8EA-6424D6A07858
Candonopsis annae (Mehes, 1914) – Higuti et al. 2009 b: 664. — Higuti et al. 2010: 267.
Diagnosis
A species of the tribe Candonini , with elongated valves, greatest height well behind the middle; carapace in lateral view with weakly rounded dorsal margin, not straight. Anterior calcified inner lamella in both valves broad; almost 1/4 of total length and with inner margin sinuous, not parallel to valve margin. Base of right prehensile palp broad, almost triangular, distal part curved, distal tip slightly expanded, set with two subequal lateral setae. Left prehensile palp larger and more elongated, with hook-like distal part,
distal tip slightly swollen. Hemipenis with triangular ls, distinctly pointed, proximo-dorsal expansion small, ms rounded and without ventral, wart-like expansion.
Etymology
The new species is named after Prof Dr Carlos Eduardo Falavigna da Rocha (USP, São Paulo, Brazil), in recognition of his vast contributions to the taxonomy of Brazilian Copepoda, but also in acknowledgement of his initiative to start research on terrestrial Ostracoda in South America.
Type material
All type material was collected on 10 Nov. 2004 by the authors, by washing roots of Eichhornia crassipes ( PAR 195) and Pistia stratiotes ( PAR 193) over a hand net with mesh size 160 µm. See Table 1 View Table 1 for measurements of water chemistry at time of collecting.
Holotype
1 ³, with soft parts dissected in glycerine in a sealed slide, and with valves stored dry ( MZUSP.28102).
Allotype
1 ♀, dissected and stored as the male ( MZUSP.28103).
Paratypes
2 ³³ dissected and stored as the holotype ( MZUSP.28104, MZUSP.28105); 4 ³ carapaces stored dry in micropalaeontological slides ( OC.3291, MZUSP.28106, MZUSP.28107, MZUSP.28108); 4 ♀♀ dissected and stored as the holotype ( OC.3292, MZUSP.28109, MZUSP.28111, MZUSP.28110); 3 ♀ carapaces stored dry in micropalaeontological slides ( OC.3293, MZUSP.28112, OC.3294).
Type locality
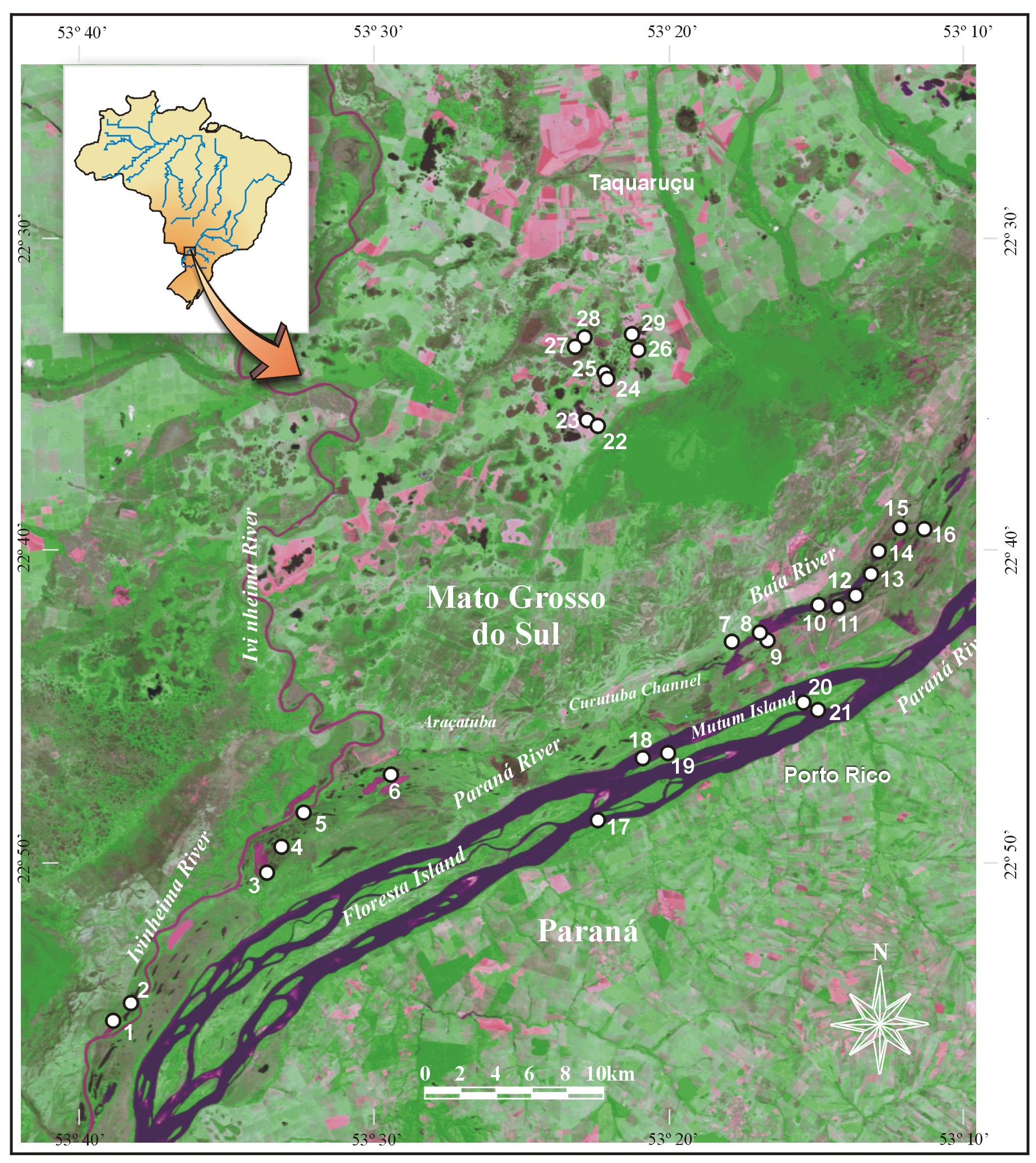
Baía River in Upper Paraná River Floodplain, in floating plants ( E. crassipes , P. stratiotes ), coordinates: 22º40’37.5” S, 53º12’29” W (locality number 13 with samples PAR193 and PAR 195 on Figure 1 View Fig ).
Other material investigated
See Table 1 View Table 1 for a list of localities where the present species was collected. Specimens MZUSP.28113, MZUSP.28114, MZUSP.28115 (³³) and MZUSP.28116 (♀) are here used for illustrations and are also deposited in the Museu de Zoologia da Universidade de São Paulo, São Paulo ( Brazil). Illustrated specimens are listed in Table 2.
Differential diagnosis
The new species differs from the congeneric species ( C. brasiliensis ), and indeed from all other Candonopsini , by the shape of the valves, by the peculiar shape of the prehensile palp (with distal part swollen in Rpp) and by the size and shape of the hemipenis.
Description of male
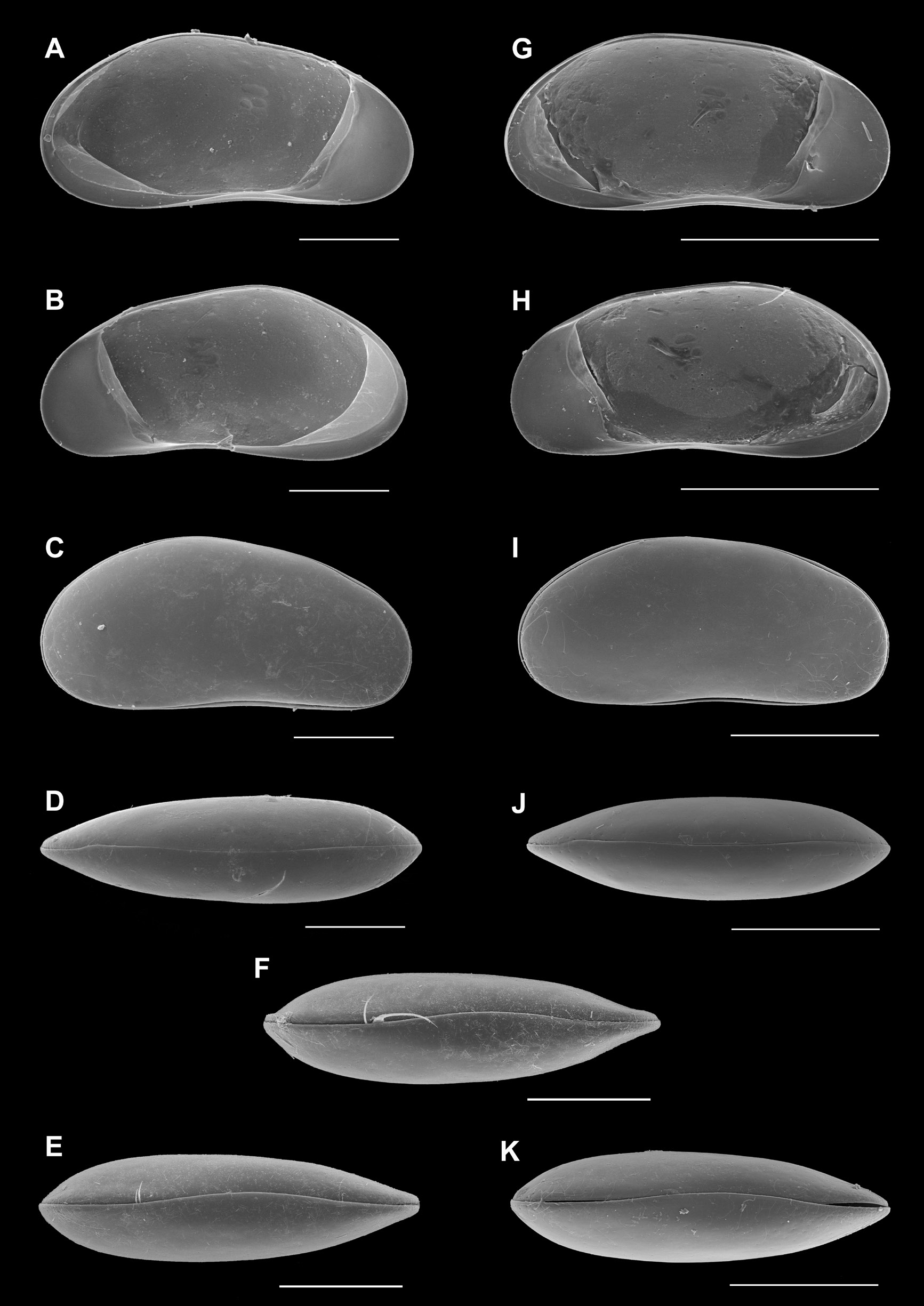
Valves ( Fig. 2A, B View Fig ) elongated, with greatest height situated behind the middle (and bluntly pointed there) anteriorly rather narrowly and posteriorly rather broadly rounded, RV and LV of highly similar shape; anterior calcified inner lamella broadly rounded, posterior calcified inner lamella very narrow and almost disappearing towards the dorsal side; inner margin of anterior calcified inner lamella slightly sinuous in both valves.
Carapace in right lateral view ( Fig. 2C View Fig ) showing LV overlapping RV slightly on all sides; external surface of valves smooth.
In dorsal view ( Fig. 2D View Fig ), carapace lancet-shaped, sharply pointed in the anteriorly, more broadly so posteriorly, greatest width situated well behind the middle.
Carapace also in ventral view ( Fig. 2E, F View Fig ) lancet-shaped, with anterior side more acutely pointed than posterior side.
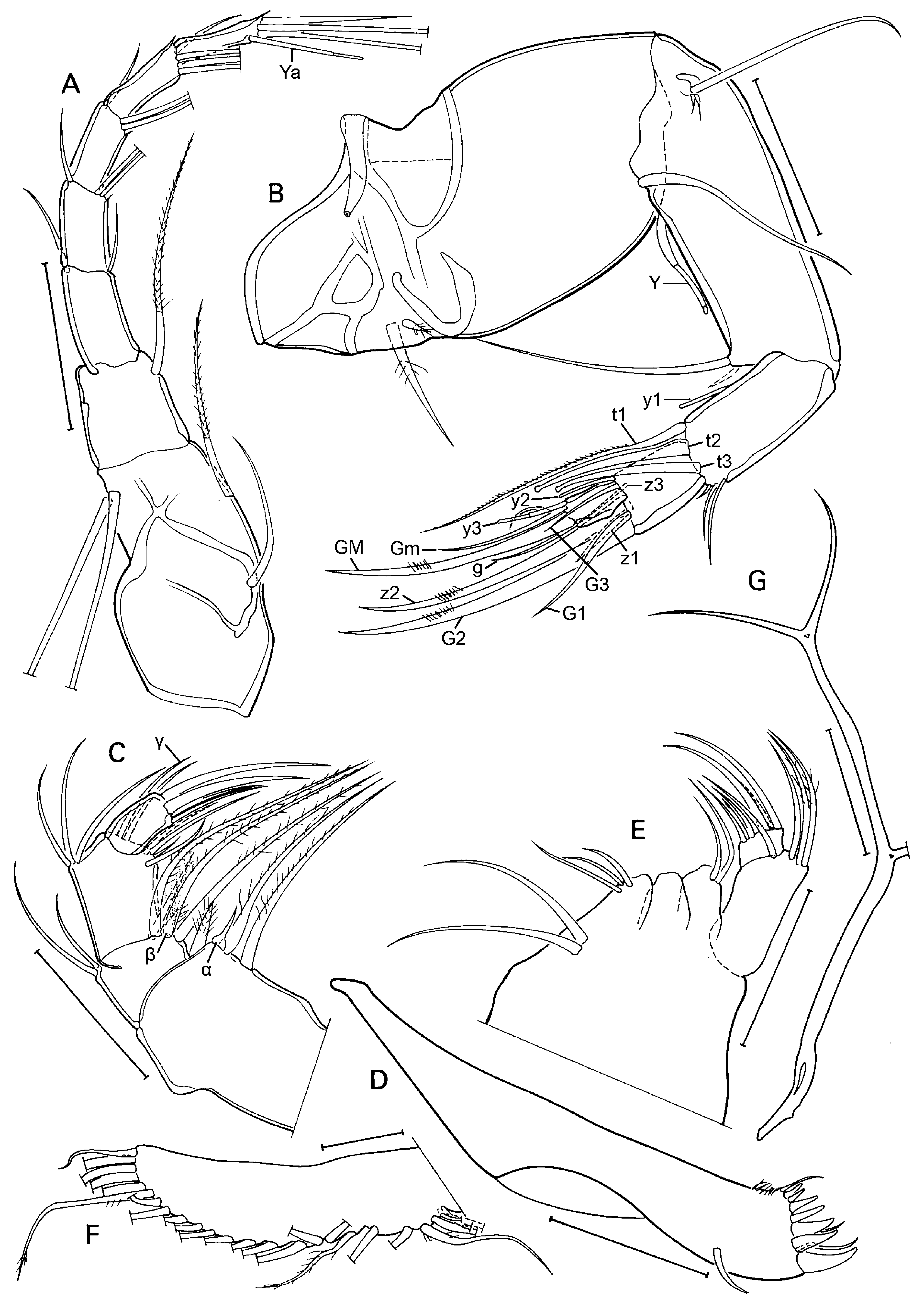
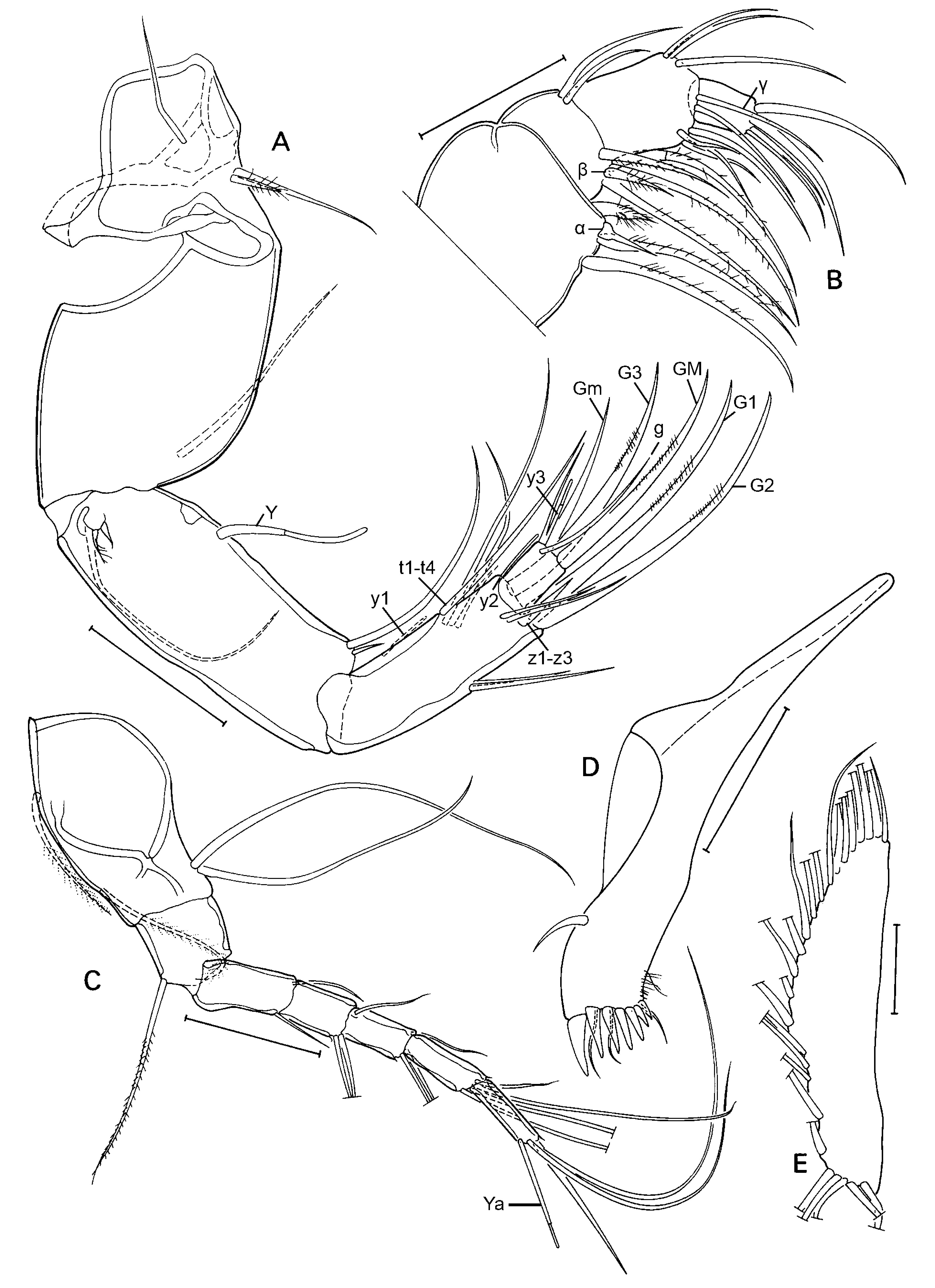
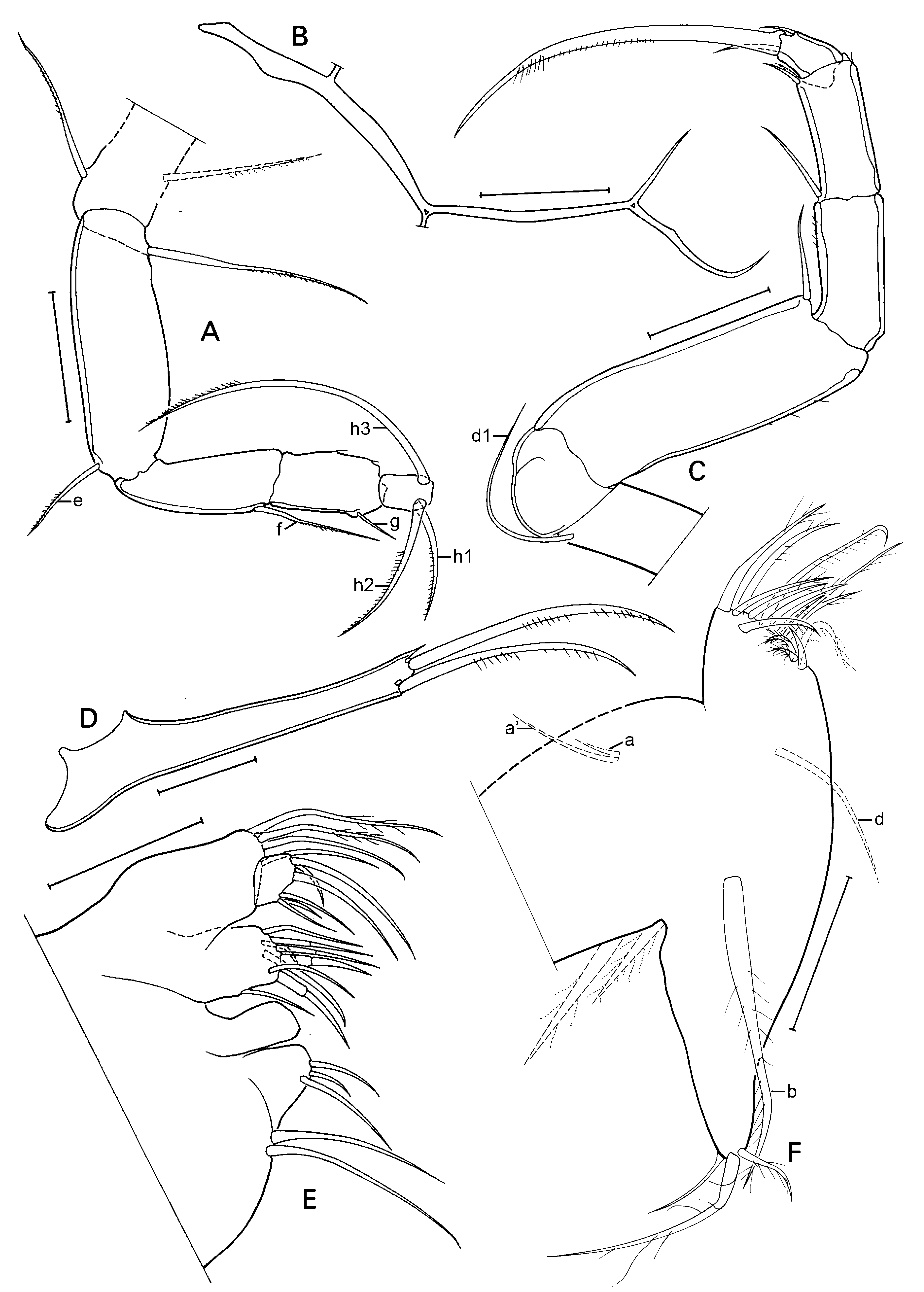
A1 ( Fig. 3A View Fig ) with 5 terminal segments relatively elongated (L = at least 1.5 x basal width). Basal part of A1 (= undivided protopodite) with 2 long sub-apical, ventral setae and two shorter setae on the dorsal side, one at ca. mid-length, the other sub-apical. Next (second) segment with one long (reaching beyond basis of penultimate segment) dorso-apical seta, no ventro-apical seta present. Third segment with one small sub-apical seta on both ventral and dorsal sides. Fourth and fifth segments with two longer dorsoapical and 1 shorter ventro-apical setae. Sixth segment with three longer dorso-apical and one shorter ventro-apical setae. Seventh (terminal segment) most elongate of all, slightly sinuous, bearing one short and two longer setae and one aesthetasc Ya, the latter shorter than the shorter seta.
A2 ( Fig. 3B View Fig ) with basal segment very broad, wide and relatively long, basally with 2 unequal setae, one relatively long and slender, the other very short and broad, both hirsute; apically with a ventral seta. Remnant of exopod consisting of a short plate, one long and two very short setae. Endopod consisting of 4 segments (penultimate segment divided). First endopodal segment long, carrying one long aesthetasc Y on the ventro-basal side, and one long and one short ventro-apical setae. Second endopodal segment shorter and smaller, but still rectangular with one ventral aesthetasc y1 inserted about mid length, 3 t-setae, with t1 a long, hirsute setae, t2 and t3 modified in the male-bristles. Dorso-apically with 3 short setae of unequal length. Third endopodal segment with apical chaetotaxy sexually dimorphic, with z1 and z3 being short setae, z2 being modified into a long claw; G2 a long claw, G1 a long seta and G3 a short setae, aesthetasc y2 short and ventro-apically inserted. Terminal (4 th) segment small, distally with claws GM (long) and Gm (short), aesthetasc y3 with its companion seta, fused at the basis and of c equal length, and seta g, slightly longer than y3.
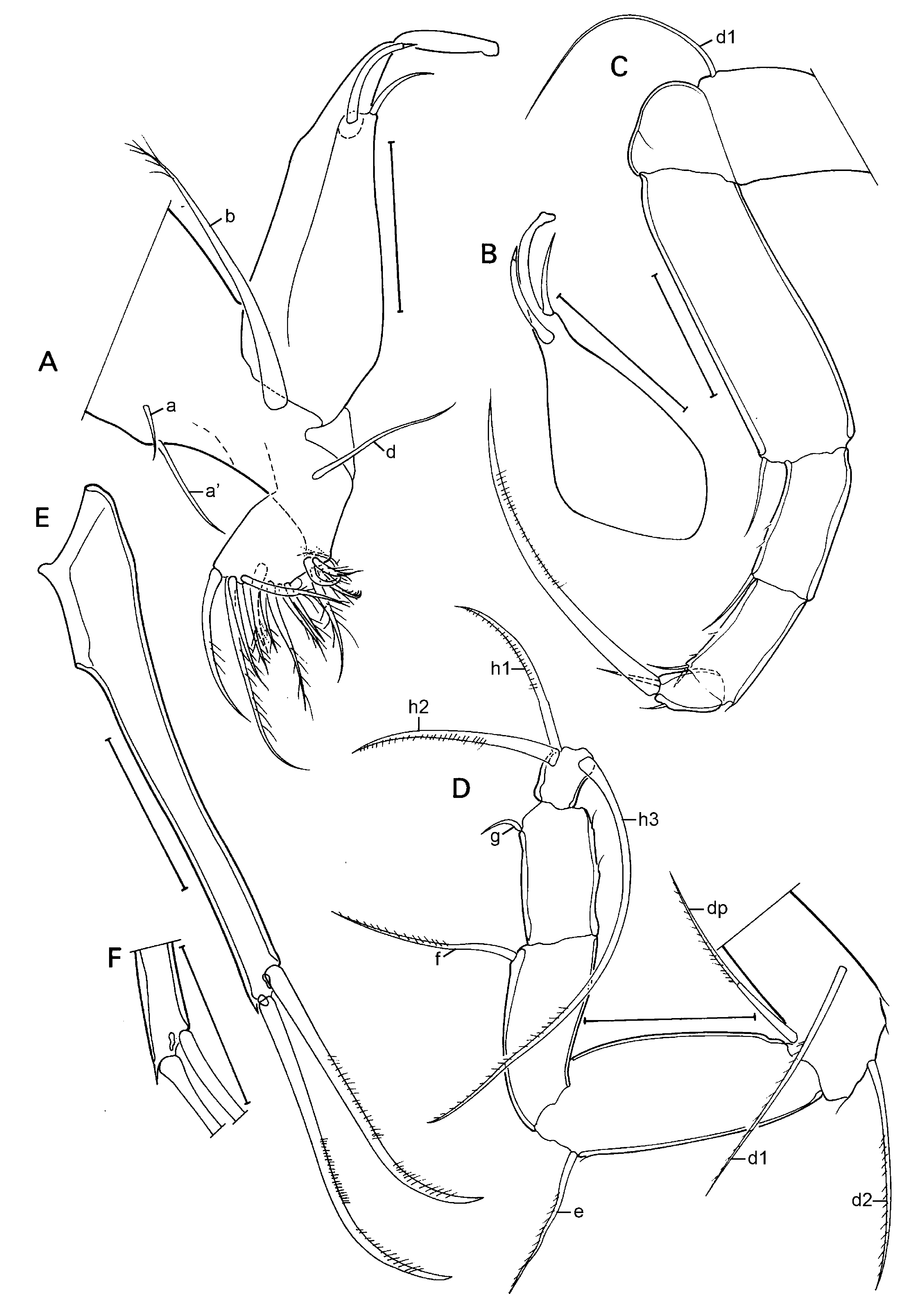
Md with coxa ( Fig. 3D View Fig ) relatively elongated, distally set with a series of strong teeth. Md-palp ( Fig. 3C View Fig ) consisting of 4 segments. Basal segment dorsally carrying the respiratory plate (not shown), ventroapically with 2 long hirsute setae (s1 & s2) and the alpha seta, the latter proximally with a broad base and a flagellum-like, distal part. Second segment dorso-apically with 2 seta of unequal length, ventro-basally with a short, stout and hirsute seta and ventro-apically with a group of 5 setae: 3 long and hirsute setae (similar to the two s-setae of the previous segment), a shorter, less hirsute seta and the short and hirsute beta-seta. Third segment with a group of 3 dorso-subapical setae, the latter smooth, a central group of two setae, one of which being the gamma seta and ventro-apical group of 4 unequal setae. Terminal segment rectangular, apically set with two large claws and an uncertain number of smaller setae.
Mx1 ( Fig. 3E View Fig ) with a basal (basipodite) part carrying a large respiratory plate (exopodite), 3 endites and a two-segmented palp (endopodite). Respiratory plate elongated, carrying more than 20 respiratory rays, some quite short, others long ( Fig. 3F View Fig ). Palp with first segment carrying 3+1 apical setae; terminal segment short and broad, carrying 2 longer claws and c 3 short setae. Chaetotaxy of three endites impossible to determine. Sideways directed bristles near first endite long and stout.
T1 (sometimes called Mx2 – Fig. 4A View Fig ) consisting of basal part (basipodite), carrying respiratory plates (not shown), a palp (modified to prehensile palp in males) and an exopodite (?) distally set with 14-15 setae of different morphology and length. Basal plate set with one long and stout ‘b’-seta, a shorter and more slender ‘d’-seta and two ‘a’-setae of unequal length. Prehensile palps ( Fig. 4A, B View Fig ) one-segmented, distally hook-like and set with two unequal, sub-apical setae; palps slightly asymmetrical, right prehensile palp ( Fig. 4B View Fig ) basally slightly more swollen and with distal hook like expansion apically swollen.
T2 (walking limb – Fig. 4C View Fig ) with 4-segmented endopodite (penultimate segment divided) and elongated. First segment with long seta d1. Knee-segment devoid of seta d2. First segment of endopod especially elongated, with short ventro-apical seta. Second segment also with one short ventro-apical seta. Third segment with two ventro-apical seta, one short, one slightly longer. Terminal segment with one short apical and one short sub-apical seta and a long apical claw.
T3 (cleaning limb – Fig. 4D View Fig ) as typical of the family. First segment with three long setae, one medial (d1), one subapical (d2) and one apical (dp). Second segment with one subapical seta (e). Third segment with one long subapical seta (f). Penultimate segment with one short and curved subapical seta (g). Terminal segment well-separated from penultimate segment and carrying three setae: one long and reflexed (h3) and two side-ways directed (h1 & h2), the latter subequal.
Caudal ramus (furca – Fig. 4E View Fig ) with stout ramus and two stout apical claws. Proximal setae missing, distal seta a small spine. Attachment to caudal ramus ( Fig. 3G View Fig ) long and stout, distally bifurcated and with additional lateral branch at c mid-length. Length ratio ramus/largest claw = 1.51.
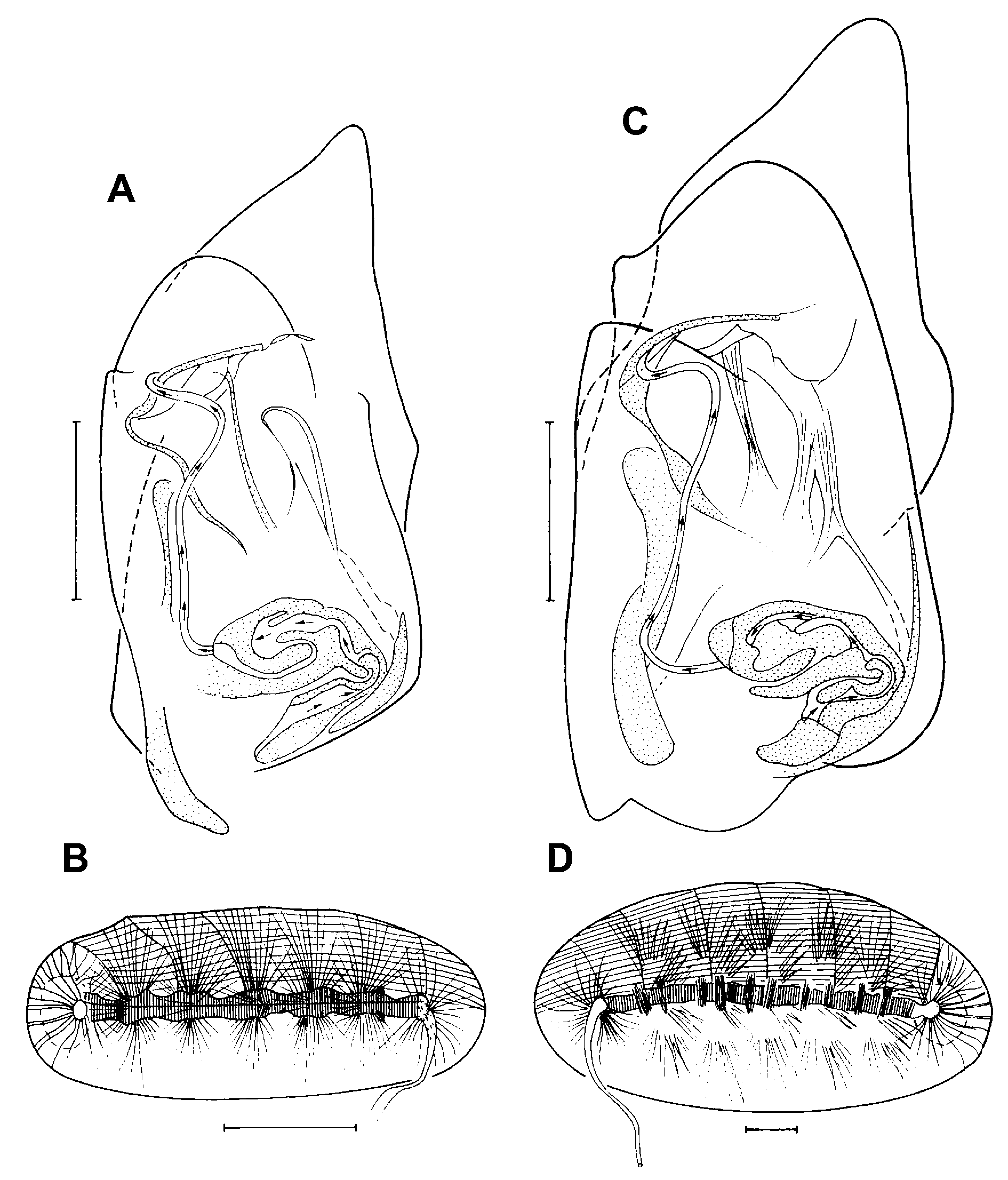
Hemipenis ( Fig. 7A View Fig ) relatively small, with rounded ms and bluntly pointed, triangular ls, the latter furthermore with small proximo-dorsal expansion; labyrinth short and stout, postlabyrinthal spermiductus narrow and straight, without additional coils, but with a weak bent at the most distal part.
Zenker organ ( Fig. 7B View Fig ) short and broad, with at least 5 spinous whirls.
Description of female
Valves ( Fig. 2G, H View Fig ) and Cp ( Fig. 2I View Fig ) in lateral view similar to those in the male, RV in inner view dorsally more rounded and less bluntly pointed than in the male. Cp in D and V views ( Fig. 2J, K View Fig ) equally narrow and lancet-shaped, in D view with slight rostrum, in ventral view without the pronounced (anterior and posterior) rostral keels, typical of the male.
Soft parts ( Figs 5 View Fig B-E, 6A-E) largely as in the male, but with sexually dimorphic A2 and T1.
A2 ( Fig. 5A View Fig ) with setae t1-4 setae like, not transformed; z1 and z2 short and slender, seta z3 short and stout. Claws G1, G2, G3 and GM all reaching to about the same point.
T1 ( Fig. 6F View Fig ) with basal part as in the male. Endopod a broad palp, with two short and one long distal setae.
Caudal ramus: length ratio ramus/largest claw = 1.40.
Measurements
See Table 3.
Ecology
See below under C. brasiliensis .
Remarks
This species was originally identified by us as Candonopsis annae (Mehes, 1914) , but subsequent detailed analysis of the valves and soft parts showed that this was not so, and that the populations from the alluvial valley of the Upper Paraná River needed be described as a new species.
Some specimens ( Fig. 2F View Fig ) give the impression that they have an anterior rostrum and a postero-ventral keel. This was observed in 2 male specimens, after their carapaces were dried for SEM. So either these 2 specimens belong to a different species, which is not very likely, or the weak calcification of the valves (typical of Candonopsis -like species) has made those parts of the valve implode while drying in air.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Candobrasilopsis rochai
| Higuti, Janet & Martens, Koen 2012 |
Candonopsis annae (Mehes, 1914)
| Higuti J. & Declerck S. A. J. & Lansac-Toha F. A. & Velho L. F. M. & Martens K. 2010: 267 |
| Higuti J. & Lansac Toha F. A. & Velho L. F. M. & Martens K. 2009: 664 |