
Calycopis bellera ( Hewitson, 1877 )
|
publication ID |
https://doi.org/10.5281/zenodo.192050 |
|
DOI |
https://doi.org/10.5281/zenodo.5627396 |
|
persistent identifier |
https://treatment.plazi.org/id/038EE54B-FFB1-FF9B-FF5C-FF09FEF4829E |
|
treatment provided by |
Plazi |
|
scientific name |
Calycopis bellera ( Hewitson, 1877 ) |
| status |
|
Calycopis bellera ( Hewitson, 1877) View in CoL ( Figs. 3–38 View FIGURES 3 – 12. C View FIGURES 13 – 18. C View FIGURES 19 – 24. C View FIGURES 25 – 26. C View FIGURE 27. C View FIGURES 28 – 30. C View FIGURES 31 – 38. C )
Egg ( Figs. 3, 4 View FIGURES 3 – 12. C , 19–24 View FIGURES 19 – 24. C ). Immediately following oviposition, egg with blue tint, but whitening as embryo develops; exochorion always whitish. Similar to C. caulonia , C. cecrops , and C. isobeon . Differing from C. caulonia in having only three or four micropylar openings and no chorionic thickenings or “islands” protruding from floor of rosette cells ( Fig. 21 View FIGURES 19 – 24. C ). Exochorion sculptured with pentagonal and hexagonal cells outlined by intersected carinate ribs ( Fig. 22 View FIGURES 19 – 24. C ). Aeropyles opening on top of elongate spinelike protuberances ( Figs. 23, 24 View FIGURES 19 – 24. C ). First instar may eat the chorion partially or entirely ( Fig. 4 View FIGURES 3 – 12. C ).
Diameter 0.76–0.86 mm (X = 0.81 mm, SD = 0.03, N = 39); height 0.40–0.50 mm (X = 0.43 mm, SD = 0.03, N = 30). Duration 6–11 days (X = 8.89 days, SD = 1.59, N = 62).
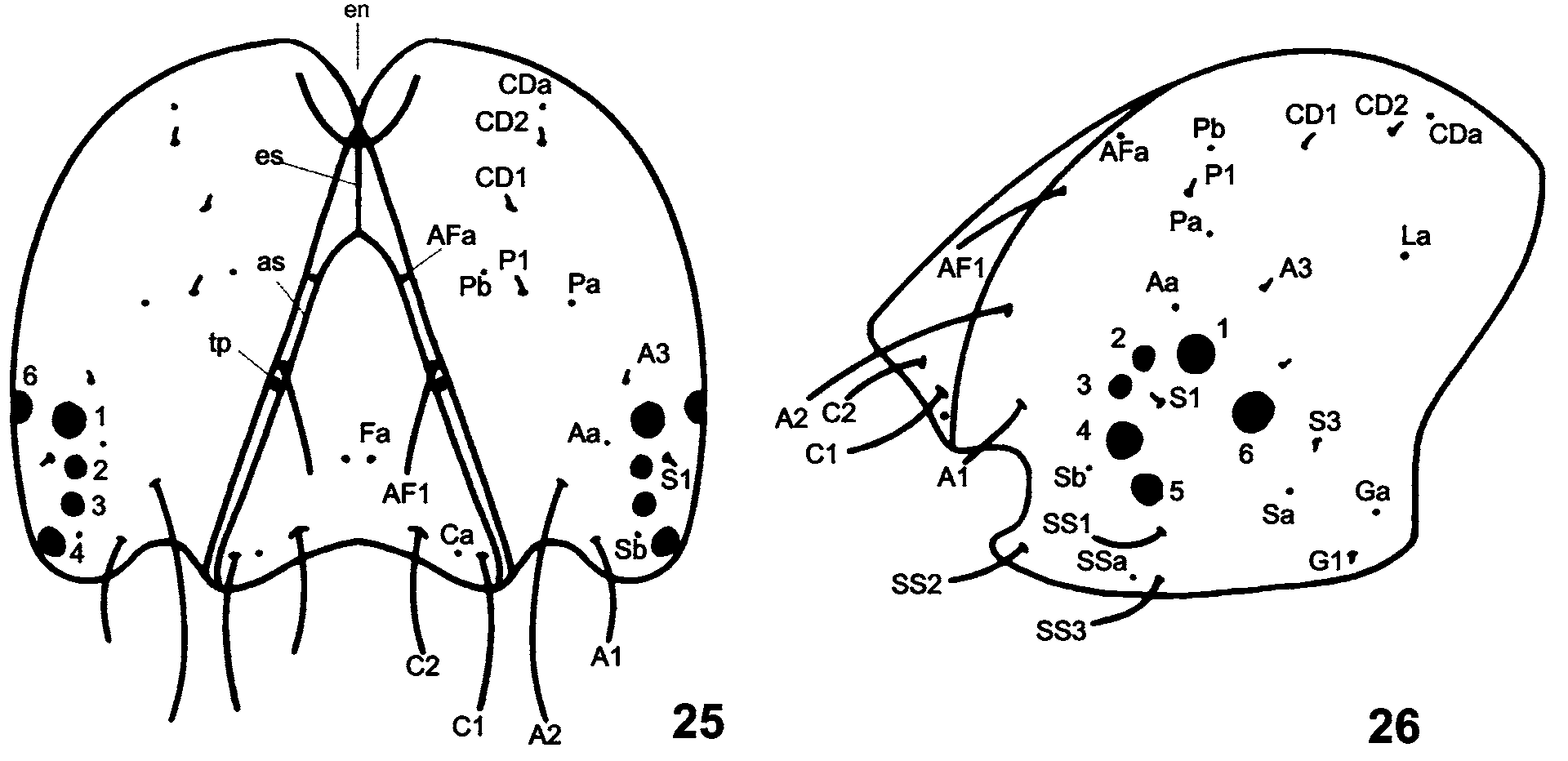
First instar ( Figs. 5, 6 View FIGURES 3 – 12. C , 25–29 View FIGURES 25 – 26. C View FIGURE 27. C View FIGURES 28 – 30. C , 31, 32 View FIGURES 31 – 38. C ). Superficially similar to other Calycopis species previously described. Head sub-quadrangular, hypognathous, uniform pale brown, with or without darker maculae; smooth surface, with prominent hyaline setae ( sensu Ballmer & Pratt 1988) of different sizes (see details on larval chaetotaxy below). Length of frontoclypeus about five times length of epicranial suture ( Fig. 25 View FIGURES 25 – 26. C ). Anteclypeus membranous. Labrum and mandibles light brown. Mandibles with cutting edge dark brown, with six teeth; differing from C. caulonia by absence of tooth on oral surface ( Fig. 29 View FIGURES 28 – 30. C ; Duarte et al. 2005: 16, fig. 16); two mandibular setae, one nearer condyle about two times length of other (we do not know yet if mandible morphology of detritivores differs significantly from that of regular folivores). Six blackish stemmata on each side of head positioned as follows: stemmata 1–5 in semicircle; stemma 5 nearer base of antenna; stemma 6 horizontally aligned with stemma 2 ( Fig. 26 View FIGURES 25 – 26. C ). Body onisciform ( Figs. 5, 6 View FIGURES 3 – 12. C ), translucent yellowish-brown at emergence, translucent yellow after feeding on artificial diet; three vivid red longitudinal stripes from mesothorax to last abdominal segment, infrequently paler on dorsal, supra-, and subspiracular areas; subspiracular stripe on seventh abdominal segment deflecting ventrally to base of proleg of tenth abdominal segment. Integument corrugated, numerous crenulations covering outer surface except ventrally. Segments A3–A6 and A10 characterized by a pair of prolegs with uniordinal crochets in uniserial mesoseries, interrupted near center by conspicuous fleshy pad; two anterior and two posterior crochets on segments A3– A6 ( Fig. 31 View FIGURES 31 – 38. C ); segment A10 with three crochets, two anterior and one posterior ( Fig. 32 View FIGURES 31 – 38. C ).
Head capsule width 0.28–0.32 mm (X = 0.31 mm, SD = 0.02, N = 30). Body length 1.9–2.6 mm (X = 2.12 mm, SD = 0.16, N = 30). Duration 4–10 days (X = 6.28 days, SD = 1.47, N = 61).
Head chaetotaxy ( Figs. 25, 26 View FIGURES 25 – 26. C ). Excluding labrum (described separately below), 16 pairs of tactile and microscopic setae and 12 pairs of pores, distributed in groups adfrontal (AF), anterior (A), cephalo-dorsal (CD), clypeal (C), frontal (F), lateral (L), microgenal (MG), posteriodorsal (P), stemmatal (S), and substemmatal (SS). Setae AF2, CD3, F1, F2, L1, and P2 absent in C. bellera . Differs from that of C. caulonia in the following: A1 about two fifths length of A2; with third anterior seta, A3, the shortest of the group, dorsal of stemma 1; CD group comprising only two setae, CD1 and CD2; S3 posterior and ventral to stemma 6; pore Sa aligned with and ventral of S3; all SS setae equal in size.
Labrum chaetotaxy. Identical to that of C. caulonia (see Duarte et al. 2005: 12, fig. 29).
Body chaetotaxy ( Fig. 27 View FIGURE 27. C ). With 128 pairs of primary setae and 22 pairs of pore cupola organs (PCOs) in the groups dorsal (D), lateral (L), microscopic dorsal (MD), microscopic subdorsal (MSD), paraproctal (PP), subdorsal (SD), ventral (V), and XD (considered “a special group developed only on the anterior margin of the prothoracic plate” by Hinton (1946: 19), and according to Stehr (1987), it should not be considered an extra seta of dorsal group), distributed as follows:
Prothorax ( Fig. 27 View FIGURE 27. C ). Identical to that of C. caulonia first instar (see Duarte et al. 2005: 13, figs. 30, 32). The term “fringe group” (F1, F2, F3) proposed by Ballmer & Wright (2008) for those setae located anteriorly and laterally to the dorsal shield on T1, which Duarte et al. (2005) treated as MSD1(?), MSD2(?) and L1, has not been adopted in the present study because the alternative terminology of Ballmer & Wright (2008) requires further comparative studies to resolve unambiguously problems of setal homology.
Mesothorax ( Fig. 27 View FIGURE 27. C ). Differing from C. caulonia in having MD1 much shorter, about a tenth length of D1. Duarte et al. (2005) referred to the seta anterior and/or dorsal to a subdorsal pore cupola organ (SDL) as MD1. With regard to the mesothorax, this interpretation is identical to that of Ballmer & Wright (2008), but in opposition to these authors, we did not observe extra subprimary dorsal setae associated with D1 and D2 on this or the following segments.
Metathorax ( Fig. 27 View FIGURE 27. C ). Identical to that of C. caulonia first instar (see Duarte et al. 2005: 15, fig. 30), including conspicuous subdorsal pore cupola organ (SDL) almost half the width of metathorax in both species.
Abdominal segment 1 ( Fig. 27 View FIGURE 27. C ). Same number of setae as in C. caulonia first instar, but with subventral PCOs lacking (SVL of Ballmer & Pratt 1992). We follow Hinton (1946) and use the term SD1 for the tactile seta anterior or antero-dorsal to the spiracle even though in recent works the same seta has been termed MSD1 referring to its ambiguous proprioceptor property (Hassenfuss & Kristensen 2003, Ballmer & Pratt 2008). In both C. bellera and C. janeirica , SD1 is associated with a subdorsal PCO (SDL).
Abdominal segment 2 ( Fig. 27 View FIGURE 27. C ). Identical to that of C. caulonia first instar (see Duarte et al. 2005: 15, fig. 30).
Abdominal segments 3-6 ( Fig. 27 View FIGURE 27. C ). Distinguished from those of C. caulonia first instar by absence of V2 (see Duarte et al. 2005: 15, fig. 30).
Abdominal segment 7 ( Fig. 27 View FIGURE 27. C ). Like C. cecrops , C. isobeon and C. caulonia , with a dorsal PCO (DL of Ballmer & Pratt 1992) joined to chalaza of D1. Differs from C. caulonia first instar by presence of a subventral PCO associated with SV1.
Abdominal segment 8 ( Fig. 27 View FIGURE 27. C ). Differing from C. caulonia in having L3 much shorter, approximately half the length of L2.
Abdominal segments 9+10 ( Fig. 27 View FIGURE 27. C ). Setal homologies for A9 and A10 are not yet well resolved and should be considered provisional (see also Ballmer & Wright 2008). Differing from C. caulonia by lacking SD2 and a pore cupola organ anteriorly associated with suranal shield, and by presence of V1 on A9. Length and position of setae similar to those of C. caulonia .
Second instar ( Figs. 7 View FIGURES 3 – 12. C , 30 View FIGURES 28 – 30. C , 33, 34 View FIGURES 31 – 38. C ). Head translucent, turning dark brown after a few minutes; stemmata blackish. Primary setae with same distribution and size patterns of first instar, including those of labrum. Secondary setae present. Mandibles with seven teeth, six visible externally, one located internally ( Fig. 30 View FIGURES 28 – 30. C ); same number of setae as first instar. Integument yellowish dorso-laterally, whitish ventrally, with two reddish longitudinal stripes extending from mesothorax to last abdominal segment: one stripe located mesodorsally; the other stripe paler and located dorsal to spiracle, consisting of oblique lines converging to base of anal proleg. Integument with brown, spiculate secondary setae, supported by conical chalazas radially distributed in nipplelike dorsal and subspiracular processes ( Fig. 7 View FIGURES 3 – 12. C ). Dentritic setae (see also Duarte et al. 2005: 17, fig. 53) interspersed with spiculate setae on this and later instars. Dendritic setae also present in C. caulonia (Hewitson) , C. isobeon (Butler & Druce) , and C. vitruvia (Hewitson) ( Duarte et al., 2005; Duarte, unpubl.). These species along with C. bellera and 15 other Calycopis species ( C. cecrops (Fabricius) , C. bactra ( Hewitson, 1877) , C. lerbela Field, 1967 , C. nicolayi Field, 1967 , C. xeneta ( Hewitson, 1877) , C. centoripa (Hewitson, 1868) , C. thama ( Hewitson, 1877) , C. torqueor (H. H. Druce, 1907) , C. vibulena ( Hewitson, 1877) , C. origo (Godman & Salvin, 1887) , C. partunda ( Hewitson, 1877) , C. indigo (H. H. Druce, 1907) , C. anfracta (H. H. Druce, 1907) , C. anastasia Field, 1967 , and C. fractunda Field, 1967 ) have been considered to constitute a natural group supported by male genitalic characters (Duarte & Robbins in preparation). Other characteristics identical to C. caulonia : prothorax wider and shorter than other thoracic segments; prothoracic shield trapezoidal, longer than wide, rounded anteriorly, number and position of setae and pore cupola organs variable; integument highly sculptured and distinctive, with uniformly-spaced oval depressions over epicuticular surface; pattern of oval depression similar in remaining instars; prolegs with uniordinal crochets in uniserial mesoseries, interrupted near center by a conspicuous fleshy pad ( Figs. 33, 34 View FIGURES 31 – 38. C ), separating the crochets into anterior and posterior groups.
Head capsule width 0.44–0.48 mm (X = 0.46 mm, SD = 0.02; N = 30). Body length 2.88–4.68 mm (X = 3.72 mm, SD = 0.58, N = 30). Duration 4–7 days (X = 5.15 days, SD = 0.66, N = 60).
Third instar ( Figs. 8 View FIGURES 3 – 12. C , 35, 36 View FIGURES 31 – 38. C ). Similar to previous instar. Differing superficially in having a greater number of spiculate and dendritic setae covering most of larval body, conferring a darker color pattern to it. Prothoracic shield and prolegs identical to those of C. caulonia ( Duarte et al. 2005: 19, figs. 33, 57).
Head capsule width 0.52–0.76 mm (X = 0.64 mm, SD = 0.06; N = 30). Maximum body length 9.20 mm (X = 8.64 mm, SD = 0.68, N = 30). Duration 5–9 days (X = 6.43 days, SD = 0.96, N = 60).
Fourth instar ( Figs. 9 View FIGURES 3 – 12. C , 37, 38 View FIGURES 31 – 38. C ). Similar to third instar, but greater number of spiculate and dendritic setae ( Fig. 9 View FIGURES 3 – 12. C ). As in C. caulonia , mandibles with three setae rather than two. Integument yellowish, but darker than in previous instar due to abundance of setae covering larval body; longitudinal stripes conspicuous, similarly positioned as second instar larva. Other morphological characteristics not differing from that described for C. caulonia .
Head capsule width 0.92–1.12 mm (X = 0.97 mm, SD = 0.06; N = 30). Maximum body length 12.64 mm (X = 11.68 mm, SD = 0.96, N = 10). Duration 4–10 days (X = 6.66 days, SD = 1.48, N = 56).
Fifth instar ( Figs. 10–12 View FIGURES 3 – 12. C ). Color pattern and shape of head, labrum, and mandibles similar to previous instar. Greater number of secondary setae, concentrated on antero-medial region of frontoclypeus. Ground color of integument yellowish or greenish ( Figs 10–12 View FIGURES 3 – 12. C ). Prolegs identical to previous instar, except for number of crochets, but with same distribution and arrangement. Other morphological characteristics as described for C. caulonia . Mature larvae of C. bellera also lack a dorsal nectar organ. However, as with other non-myrmecophilous lycaenid larvae, they possess PCOs, which are hypothesized to release substances that deter aggression in ants. Larvae of Calycopis have not been reported being tended by ants either in the laboratory or in nature, as in some eumaeines ( Pierce et al. 2002). In addition to PCOs, we also found specialized setae (dendritic setae of Ballmer & Pratt 1988) considered to be important in maintaining communication between larvae and ants (see Fiedler 1991; Pierce et al. 2002).
Head capsule width 1.28–1.44 mm (X = 1.38 mm, SD = 0.06; N = 30). Maximum body length 17.60 mm (X = 14.25 mm, SD = 1.98, N = 30). Duration 6–20 days (X = 12.21 days, SD = 2.80, N = 53).
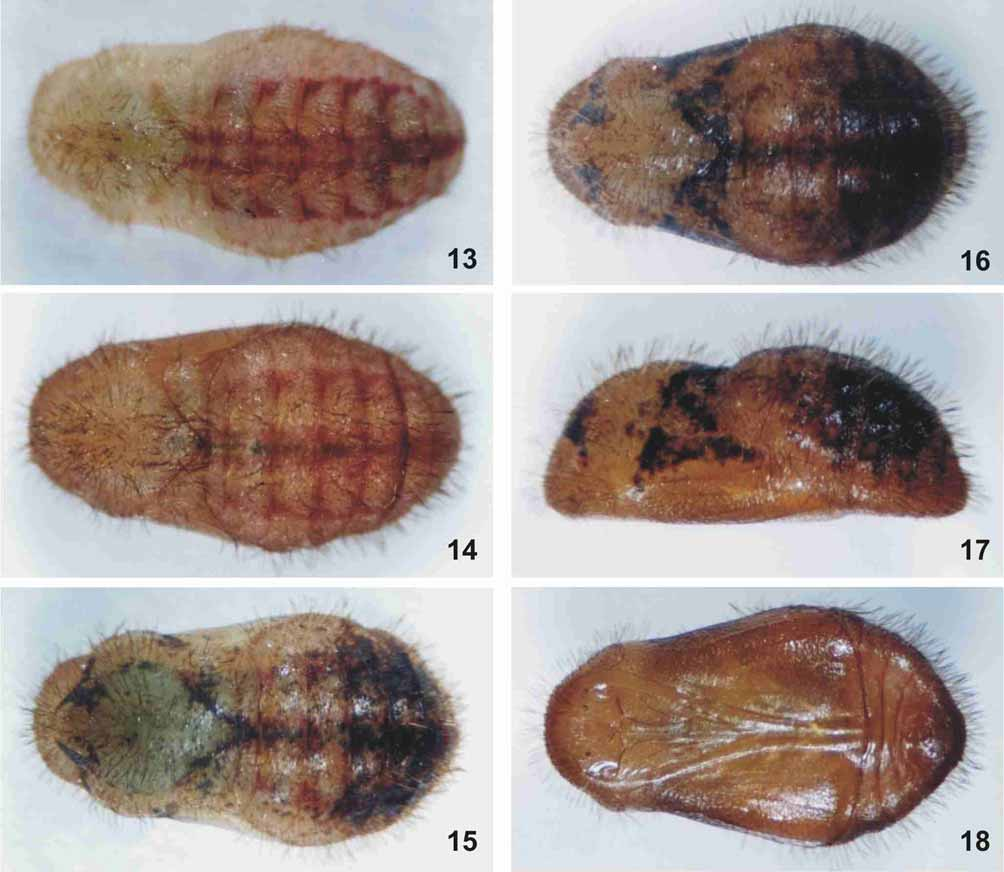
Pupa ( Figs. 13–18 View FIGURES 13 – 18. C ). Identical to that of C. caulonia . Head and thorax initially translucent light green ( Fig. 13 View FIGURES 13 – 18. C ). Abdomen dorsally with vivid red maculae extending laterally, but not reaching spiracles ( Fig. 13 View FIGURES 13 – 18. C ). Integument darkening gradually in some individuals ( Figs. 14–18 View FIGURES 13 – 18. C ). Dark brown maculae scattered on dorsal and lateral body, densely covered with small golden, prominent setae.
Pupal width on metathorax 3.0–4.0 mm (X = 3.2 mm, SD = 0.3, N = 30), on segment A3 4.0–6.0 mm (X = 4.8 mm, SD = 0.4, N = 30). Pupal length 7.9–11.1 mm (X = 9.0 mm, SD = 0.8, N = 30). Duration 11–17 days (X = 12.56 days, SD = 1.16, N = 41).
Larval development on artificial diet. All five larval instars fed on artificial diet, which supported complete development, as in C. caulonia . Data on development times from egg to adult are summarized in Table 1 View TABLE 1 . Females of C. bellera emerged from pupae as adults earlier on average (X = 55.28 days, SD = 4.21, N = 25) than males (X = 59.13 days, SD = 3.69, N = 16).
Cannibalism. When confined in containers, cannibalism among mature larvae was frequent as it was in C. caulonia . Cannibalistic behavior is likely to be widespread in the genus.
Duration (days) Range
X SD
Species Interval (days) N Egg to adult 56.78 4.40 50–68 41 Taxonomy. Reared females ( Fig. 1 View FIGURE 1. C b) match the female lectotype of Thecla bellera deposited in the Natural History Museum (London), which was designated by Field (1967a). The dorsal surface is brown without shining blue scales, which differentiates this species from related Calycopis species. Reared males ( Fig. 1 View FIGURE 1. C a) match the male holotype of Thecla devia deposited in Museum für Naturkunde der Humboldt- Universität (Berlin), which has a 1994 red holotype label added by G. Lamas. The ventral hindwing cubital spot is brown/black and the ventral wings lack central dark patches, which differentiates this species from other related Calycopis species ( Field 1967a). The male associated with the female in Field (1967a) has an orange-red cubital spot on the ventral hindwing and is a different species, but we are uncertain which species it represents.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Theclinae |
|
Tribe |
Eumaeini |
|
Genus |