
Calliax michelottii (A. Milne Edwards, 1860 ) Hyžný & Gašparič, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3821.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:3F7440FB-B9A6-4669-A1B2-4DAB6CFEB6B7 |
|
persistent identifier |
https://treatment.plazi.org/id/03CA87CA-FFC6-C817-00A2-FEE5FBF2B745 |
|
treatment provided by |
Felipe |
|
scientific name |
Calliax michelottii (A. Milne Edwards, 1860 ) |
| status |
comb. nov. |
Calliax michelottii (A. Milne Edwards, 1860) comb. nov.
( Figs 5A–D View FIGURE 5 , 6A–L View FIGURE 6 , 7A–M View FIGURE 7 , 8A–L View FIGURE 8 , 9A–I View FIGURE 9 , 10A–B View FIGURE 10 )
Callianassa Michelotti A. Milne Edwards, 1860: 341 View in CoL , pl. 14, fig. 3; A. Milne Edwards, 1860: 210, pl. 14, fig. 3.
Callianassa michelotti — Schweitzer et al., 2010: 35.
Callianassa Michelottii View in CoL — Fritsch, 1871: 691, pl. 17, figs. 5–13; Noetling, 1886: 84, pl. 5, fig. 4; Crema, 1895: 667, fig. 3; Glaessner, 1928: 167–168; Glaessner, 1929: 84.
Callianassa michelotii — Beurlen, 1931: 111–112.
“ Callianassa View in CoL ” michelotti — Polkowsky, 2005: 17, figs. 8–13, pl. 1, figs. 1–8.
Callianassa cf. michelottii — Moths & Montag, 2002: 7, pl. 4, fig. 3.
ʻCallianassaʼ szobensis Müller, 1984: 53 (partim), pl. 7, figs.3–4.
Callianassa szobensis — Schweitzer et al., 2010, 37.
„ Callianassa “ szobensis — Hyžný, 2011: table 2.
Callianassa sp. —Houša in Špinar et al., 1965: 734, figs. X-184–185.
? Calianassa Michelotti — Wagner-Klett, 1919: 107, pl. 2, fig. 1.
? ʻCallianassaʼ szobensis Müller, 1984: 53 (partim); pl. 7, figs. 5–6.
? Callianassa sp. — Philippe & Secretan, 1971: 128, pl. C, figs. 13–14.
? Eine nicht bestimmbare Hand— Lőrenthey, 1907: 212, 222, pl. 1, fig. 5.
non Callianassa cf. michelottii Müller, 1993: 7 , fig. 3A.
Diagnosis. Major cheliped massive; merus longer than high (L/H=2.2) with convex upper margin, carpus higher than long (L/H=0.45), about half the length of manus or shorter, upper margin straight, proximolower margin regularly rounded and smooth in outline; propodus rectangular, longer than high, exhibiting two morphotypes (L/ H=1.1–1.2; 1.3–1.5); outer lateral surface of manus smooth, adorned with several tubercles; fixed finger shorter than manus, triangular, with two parallel ridges extending onto manus, lower (less developed) ridge positioned close to the lower propodal margin; occlusal margin of fixed finger serrated and adorned with a blunt tooth pointing up and forward; dactylus slender, with serrated occlusal margin and pointed tip.
Description. Major cheliped massive. Merus longer than high (L/H=2.2), with prominent keel along the midline; upper margin unarmed, convex; lower margin concave, serrated. Carpus higher than long (L/H=0.45), upper margin straight or concave, proximolower margin regularly rounded and smooth in outline. Propodus rectangular, longer than high (L/H=1.1–1.5), distinctly longer than carpus; upper and lower margins straight, parallel to each other, forming distinct keels on the inner sides, rounded proximally; distal margin straight, parallel to proximal margin, the base of the fixed finger may form a notch. Outer lateral surface of manus smooth, adorned with several tubercles (at the bases of setal pores) lying in a row positioned obliquely. Inner lateral surface of manus concave, with distinct depression at the base of fixed finger. Fixed finger shorter than manus, triangular, with two ridges extending onto manus, lower (less developed) ridge positioned close to the lower propodal margin, upper one (strongly developed) accompanied with several tubercles proximally; occlusal margin serrated and adorned with a blunt tooth pointing up and forward. Dactylus slender, with serrated occlusal margin and pointed tip.
Variations. Major propodi of Calliax michelottii comb. nov. exhibit a certain degree of variation. When examining a large number of specimens, one can observe differences in the manus length/height ratio attaining values in two intervals, i.e. 1.1–1.2 (shorter morph) and 1.3–1.5 (longer morph). Unfortunately it is difficult to state whether it is intraspecific variation or if it mirrors sexual dimorphism. Polkowsky (2005) tentatively interpreted two morphotypes of C. michelottii as sexual dimorphs. Some propodi may possess a relatively well developed notch at the base of the fixed finger. Its development seems to be at least partly correlated with the size, but as already pointed out by Polkowsky (2005), the notch in the longer morphotype is usually better developed.
Development of the ridges is a stable character, i.e. there are always two ridges present, and the lower one is rather faint. What differs in various propodi is the shape of the upper ridge in its proximalmost part. In some specimens it is straight, but in others it is curved downwards. The development of tubercles accompanying the upper ridge also varies, depending on the size of the animal.
Glaessner (1928) reported strong ornamentation on the isolated propodi from Jarenina, Slovenia ( Figs 8A–B View FIGURE 8 ). This can, however, be ascribed mainly to preservational aspects. Specimens from Želiezovce, Slovakia ( Špinar et al. 1965: fig. X-184; refigured here as Fig. 8L View FIGURE 8 ) and Szob, Hungary ( Müller 1984: pl. 7, figs. 3, 4; refigured here as Figs 8J–K View FIGURE 8 ) exhibit a similar pattern of ornamentation, although not so pronounced as in the material from Jarenina. Possibly it is related to calcification of the cuticle. Experimental data, which are lacking at this point of time, would help in elucidating this issue.
Major P1carpus may have a concave upper margin ( Fritsch 1871: pl. 17, fig. 9; refigured here as Fig. 6E View FIGURE 6 ). A similarly concave upper margin was also observed in extant Calliax doerjesti ( Sakai 1999: fig. 28, 29; Fig. 3A View FIGURE 3 ). This feature may be related to age and/or size of the individual. For resolving this issue in Calliax michelottii comb. nov. more preserved carpi must be examined.
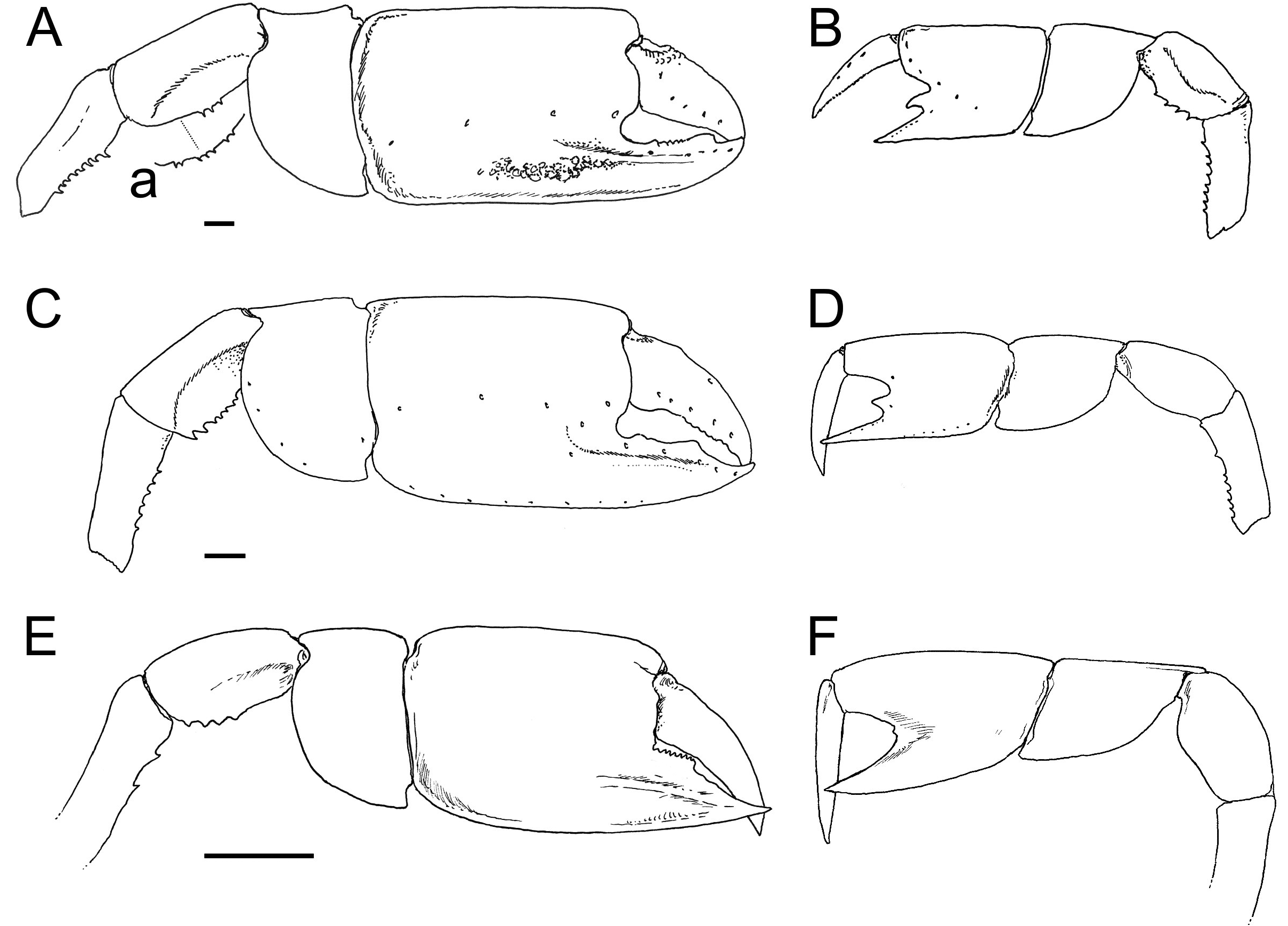
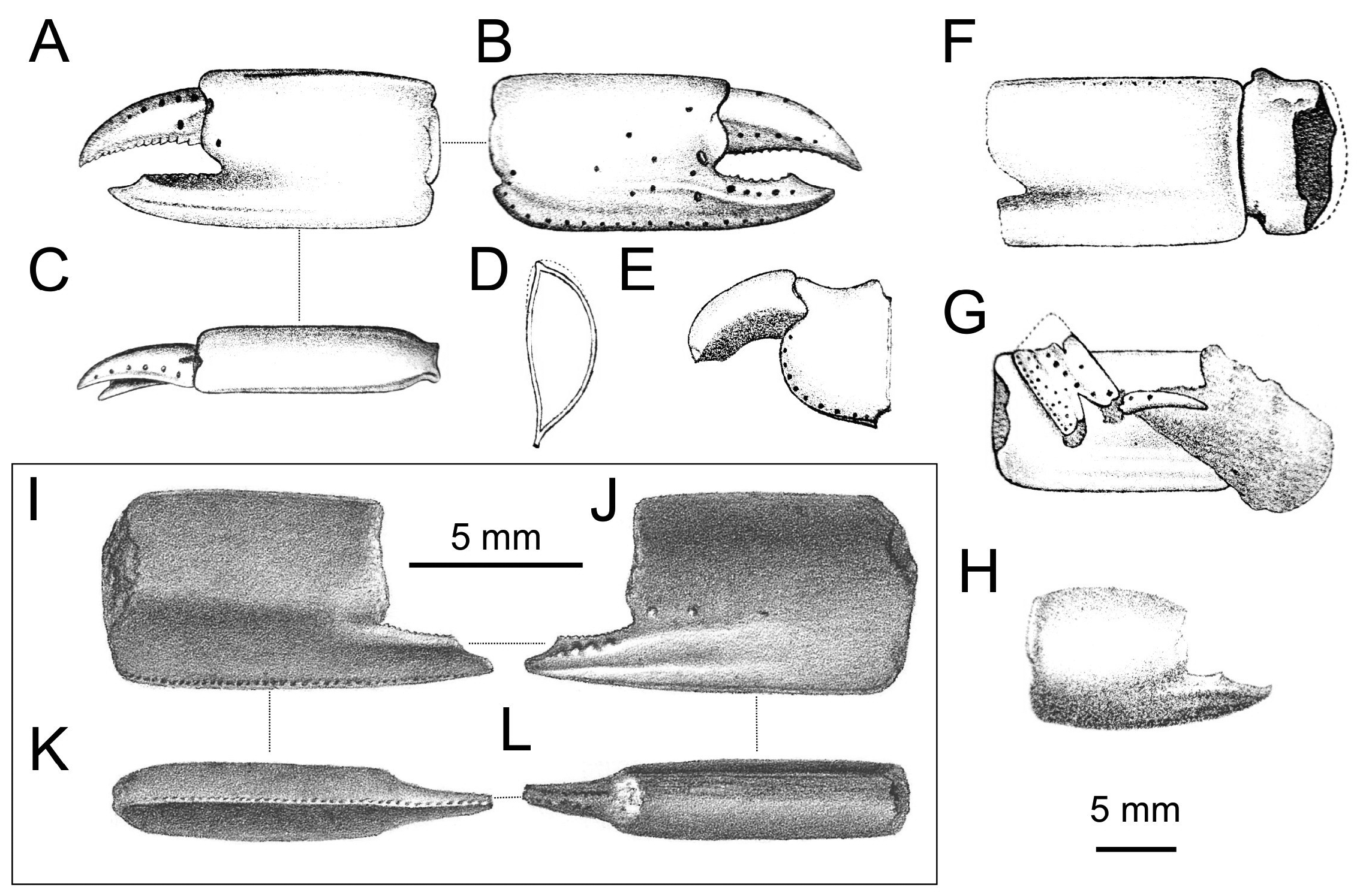
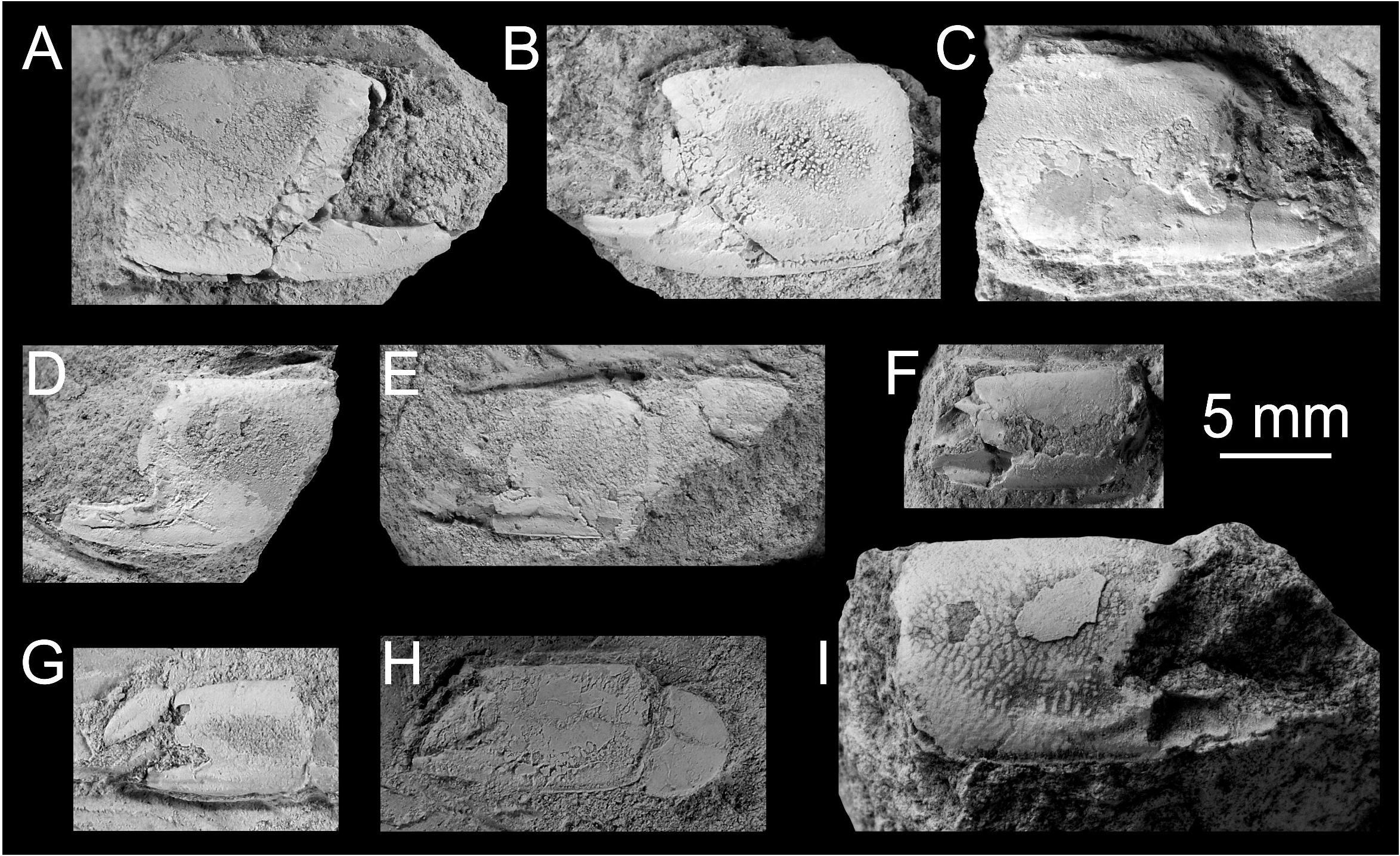
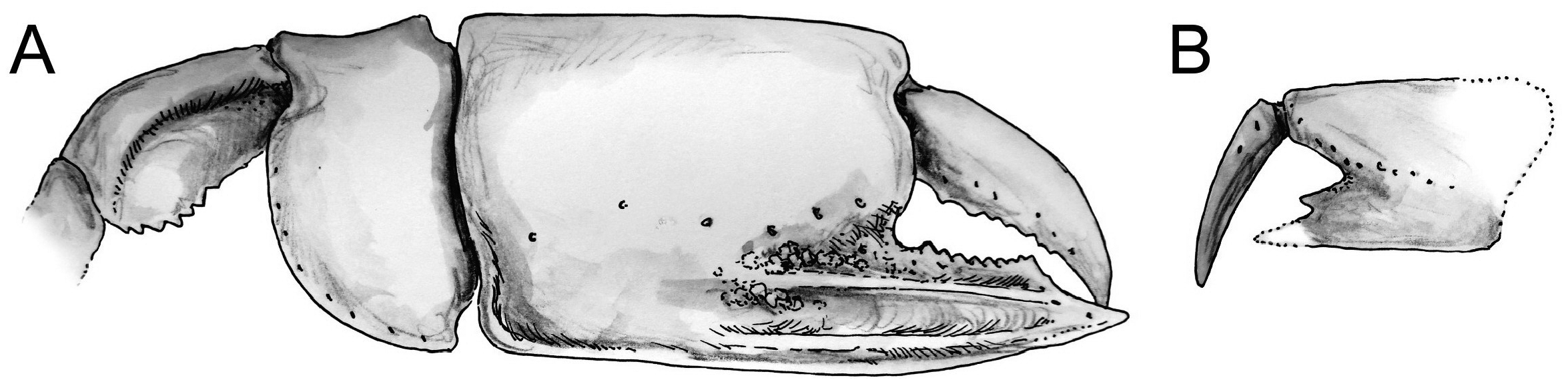
Material examined. Only remains of major chelipeds have been examined: the holotype of Callianassa michelottii from Superga at Turin, Italy (MNHN-F-B32690; Fig. 5 View FIGURE 5 ); 21 fragmentary propodi and one carpus ( NHMW 1874/0029/1154 a–v) from Sternberg, Germany ( Fig. 7 View FIGURE 7 ); one isolated propodus from Pinnow bei Schwerin, Germany ( NHMW 1874/0029/1155; Fig. 7I View FIGURE 7 ); one right propodus articulated with dactylus from Pucking, Austria ( NHMW 2003/0026/0913; Fig. 8C View FIGURE 8 ); entire articulated right cheliped from Neuhofen bei Tettenweis ( NHMW 2010/0089/0001; Fig. 8I View FIGURE 8 ); two isolated left propodi ( RGA / SMNH 0773, 0779), two isolated right propodi ( RGA / SMNH 0754, 0864), four articulated chelae consisting of propodus and dactylus ( RGA / SMNH 1075, 1187) and even carpus and merus ( RGA / SMNH 1069, 1191) from Kamnik-Košiše, Slovenia ( Figs 9A–H View FIGURE 9 ); right isolated propodus from the unknown site at Kamnik (Stein in Krain), Slovenia ( GBA 2009/014/0027; Fig. 9I View FIGURE 9 ); two isolated propodi from Jarenina (Jahring), Slovenia already reported (but not figured) by Glaessner (1928) ( UMJGP 77873–77874; Figs 8A–B View FIGURE 8 ); one right fragmentary propodus from Rohožník, Slovakia ( KGP-MH RO- 001; Figs 8D–H View FIGURE 8 ); holotype of Callianassa szobensis from Szob, Hungary (M.2004.158.1; Figs 8J–K View FIGURE 8 ).
Occurrence. Oligocene (Rupelian)–Middle Miocene (Serravallian) of Europe. The oldest occurrence of the species is known from the Rupelian of the Mainz Basin, Germany ( Fritsch 1871; Figs 6A–G View FIGURE 6 ). The youngest one is from the Middle–Late Badenian (Serravallian) of Slovakia (herein). All confirmed occurrences are summarized in Table 2 View TABLE 2 . For details see text below.
Remarks. Callianassa michelottii was originally described based on isolated propodi ( Fig. 5 View FIGURE 5 ) from the Miocene of Superga hill near Turin, Italy (A. Milne Edwards 1860). A. Milne Edwards (1860: p. 211) noted that the presence of the keel on the fixed finger accompanied by the tubercles above it is unique not only for fossil species, but this character distinguishes the species from all extant callianassids known at that time as well. Indeed, the type species of Calliax was described more than a century later by de Gaillande & Lagardère (1966). Crema (1895) reported C. michelottii from the Middle Miocene of nearby localities in the Turin hills.
Fritsch (1871) reported C. michelottii from the “Middle” Oligocene (Rupelian) “Septarienthon” of the Mainz Basin (W Germany). Interestingly, Fritsch (1871: pl. 17, figs 5–13) reported and also figured a small chela preserved together with the major P1 propodus ( Fig. 6G View FIGURE 6 ). He tentatively interpreted it as remains of P2, but it in fact represents a minor P1 chela. The description of Fritsch (1871: 696) is very clear in this sense (“Der bewegliche Finger steht auf einem weit nach vorn vorspringenden Theil des Ballens. Vom unbeweglichen Finger ist nu rein ganz geringer Theil sichtbar, warscheinlich war derselbe indess kurz, die hervortretenden Leisten desselben Gliedes der Vorderhand fehlen.” English translation: Dactylus is located on the manus portion projecting forward. Only very small portion of the index is visible. It was probably short, although the tip is missing.) Fritsch (1871: 692) mentioned also the presence of the cephalothorax (“Wohl liegt ein Exemplar vor, an welchem man Theile des kleinen dünnschaligen Cephalothorax und der seitlich zusammengedrückten hinteren Fusspaare erkennt, doch is dasselbe für eine Charakteristik des Thieres zu ungenügend erhalten.” English translation: There is a sample in which the individual shows the preserved small cephalothorax with thin cuticle and the laterally compressed “rear” pereiopod. However, the preservation of both characters is insufficient.), but he did not figure it. The material of Fritsch (1871) was reexamined by Beurlen (1931); he confirmed much of what was already done in the original work.
Noetling (1886: pl. 5, fig. 4; refigured here as Figs 6H–K View FIGURE 6 ) reported the species from the Late Oligocene “Sternberger Gestein”of Kobrow, Germany. The latest contribution on Callianassa michelottii from this facies is that by Polkowsky (2005) who discussed at length the variation of the species as well as its geographic distribution.
Lőrenthey (1907: pl. 1, fig. 5; refigured in Fig. 6L View FIGURE 6 ) reported and figured undetermined isolated propodi from the Miocene of Cagliari, Sardinia. With two parallel ridges on the fixed finger, the specimen shows affinities to Calliax michelottii comb. nov., but possesses rather convex upper propodal margin, which is unusual for Calliax , although the character itself may reflect mere intraspecific variation. The material (collection of Lovisato) is considered lost ( De Angeli & Garassino 2006); thus, the re-examination of the material is not possible. As a consequence, we treat the occurrence as questionable.
Wagner-Klett (1919) reported Callianassa michelottii from the Oligocene Septarienton of Wiesloch, Germany. The figure he provided ( Wagner-Klett 1919: pl. 2, fig. 1), however, does not conform to the diagnosis of the species. The specimen consisting of an articulated carpus, propodus, and dactylus shows no ridges on the fixed finger. The material was either not properly figured or it represents a different species.
Glaessner (1928) reported the species from the Helvetian (Early Miocene) of Jahring (today's Jarenina, Slovenia) in the Styrian Basin. He, however, did not provide figures. The re-examination of the material by the senior author confirms the identification of the material ( Figs 8A–B View FIGURE 8 ).
Beurlen (1939: 143) when discussing affinities of Callianassa brevimanus Beurlen, 1939 , (currently recognized as a member of Lepidophthalmus Holmes, 1904 ; see Hyžný & Dulai in press) briefly mentioned C. michelottii as coming from the Oligocene of the Mainz Basin and Miocene of Italy and the Vienna Basin, however, without any further reference. It is supposed here that Beurlen (1939) mentioned the occurrence from the Vienna Basin erroneously, as there is no such published report known to the authors. He might have been referring to the published occurrence from the Styrian Basin by Glaessner (1928).
Several chelae from the Middle Miocene (“Badenian”) of Želiezovce (Slovak part of the Danube Basin) figured by Houša in Špinar et al. (1965: figs. X-184–185, refigured herein as 8L) as Callianassa sp. can be clearly assigned to Callianassa michelottii . Unfortunately, the repeated search for the material was unsuccessful, thus, it is herein considered lost.
Philippe & Secretan (1971) reported 10 fragmentary propodi from the Burdigalian of SE France. The figured specimens (Pl. C, figs. 13, 14) show the keeled fixed finger accompanied with furrows, thus, pointing to attribution of the material to Calliax . The age and location of the specimens would speak for indentification as C. michelottii . Without personal re-examination of the material, however, we are reluctant to treat the specimens as conspecific with C. michelottii .
The description and figures of “ Callianassa ” szobensis Müller, 1984 from the Middle Miocene (Badenian) of Hungary fit the diagnosis and variations of C. michelottii . The propodus is longer than high, the fixed finger possesses two ridges parallel to one another, and there are tubercles on the lateral surface of the propodus ( Figs 8J–K View FIGURE 8 ). As a consequence, “ C. ” szobensis is considered a junior subjective synonym of C. michelottii .
From the Miocene of Spain (Catalonia), Müller (1993: fig. 3A) reported and figured an isolated propodus of a callianassid shrimp. He classified it as Callianassa cf. michelottii . The specimen, however, is rather dissimilar to C. michelottii ; it does not possess a double ridge on the fixed finger and has the upper margin converging proximally, which is very atypical for the latter species.
Calliax michelottii n. comb. is morphologically very close to its extant congeners. The upper margin of the major P1 merus is, however, more convex in C. michelottii n. comb. ( Fritsch 1871: pl. 17, fig. 9; see also Figs. 8I View FIGURE 8 ), whereas it is rather straight in both extant species, C. doerjesti and C. lobata ( Fig. 3A View FIGURE 3 and Fig. 3C View FIGURE 3 , respectively). The development of spines on the lower margin of the major P1 merus in Calliax michelottii comb. nov. is closer to that of C. lobata . The tooth formula of the occlusal margin of the fixed finger looks different in all three species, but this may be a matter of variation and is not considered taxonomically important here.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Axiidea |
|
Family |
|
|
SubFamily |
Eucalliacinae |
|
Genus |
Calliax michelottii (A. Milne Edwards, 1860 )
| Hyžný, Matúš & Gašparič, Rok 2014 |
Callianassa michelotti
| Schweitzer, C. E. & Feldmann, R. M. & Garassino, A. & Karasawa, H. & Schweigert, G. 2010: 35 |
Callianassa
| Polkowsky, S. 2005: 17 |
Callianassa cf. michelottii
| Moths, H. & Montag, A. 2002: 7 |
Callianassa cf. michelottii Müller, 1993: 7
| Muller, P. 1993: 7 |
szobensis Müller, 1984: 53
| Muller, P. 1984: 53 |
Callianassa sp.
| Philippe, M. & Secretan, S. 1971: 128 |
Callianassa sp.
| Spinar, Z. & Cepek, P. & Boucek, B. & Eliasova-Frajova, H. & Havlicek, V. & Horny, R. & Housa, V. & Kalabis, V. & Kukalova, J. & Marek, L. & Nekvasilova, O. & Pokorny, V. & Prokop, R. & Pribyl, A. & Ruzicka, B. & Snajdr, M. & Stemprokova, D. & Zikmundova, J. 1965: 734 |
Callianassa michelotii
| Beurlen, K. 1931: 111 |
Calianassa
| Wagner-Klett, W. 1919: 107 |
Callianassa Michelottii
| Glaessner, M. F. 1929: 84 |
| Glaessner, M. F. 1928: 167 |
| Crema, C. 1895: 667 |
| Noetling, F. 1886: 84 |
| Fritsch, K. von 1871: 691 |
Callianassa Michelotti A. Milne Edwards, 1860: 341
| Milne Edwards, A. 1860: 341 |
| Milne Edwards, A. 1860: 210 |