
Brueelia terpsichore, Gustafsson & Zou & Oslejskova & Najer & Sychra, 2019
|
publication ID |
https://doi.org/ 10.5852/ejt.2019.507 |
|
publication LSID |
lsid:zoobank.org:pub:39A9499C-7551-4821-9C1D-4FA3BA0AD533 |
|
DOI |
https://doi.org/10.5281/zenodo.5628144 |
|
persistent identifier |
https://treatment.plazi.org/id/A5296B28-CC4A-430B-928B-9A92B2AC5E2B |
|
taxon LSID |
lsid:zoobank.org:act:A5296B28-CC4A-430B-928B-9A92B2AC5E2B |
|
treatment provided by |
Plazi |
|
scientific name |
Brueelia terpsichore |
| status |
sp. nov. |
Brueelia terpsichore sp. nov.
urn:lsid:zoobank.org:act:A5296B28-CC4A-430B-928B-9A92B2AC5E2B
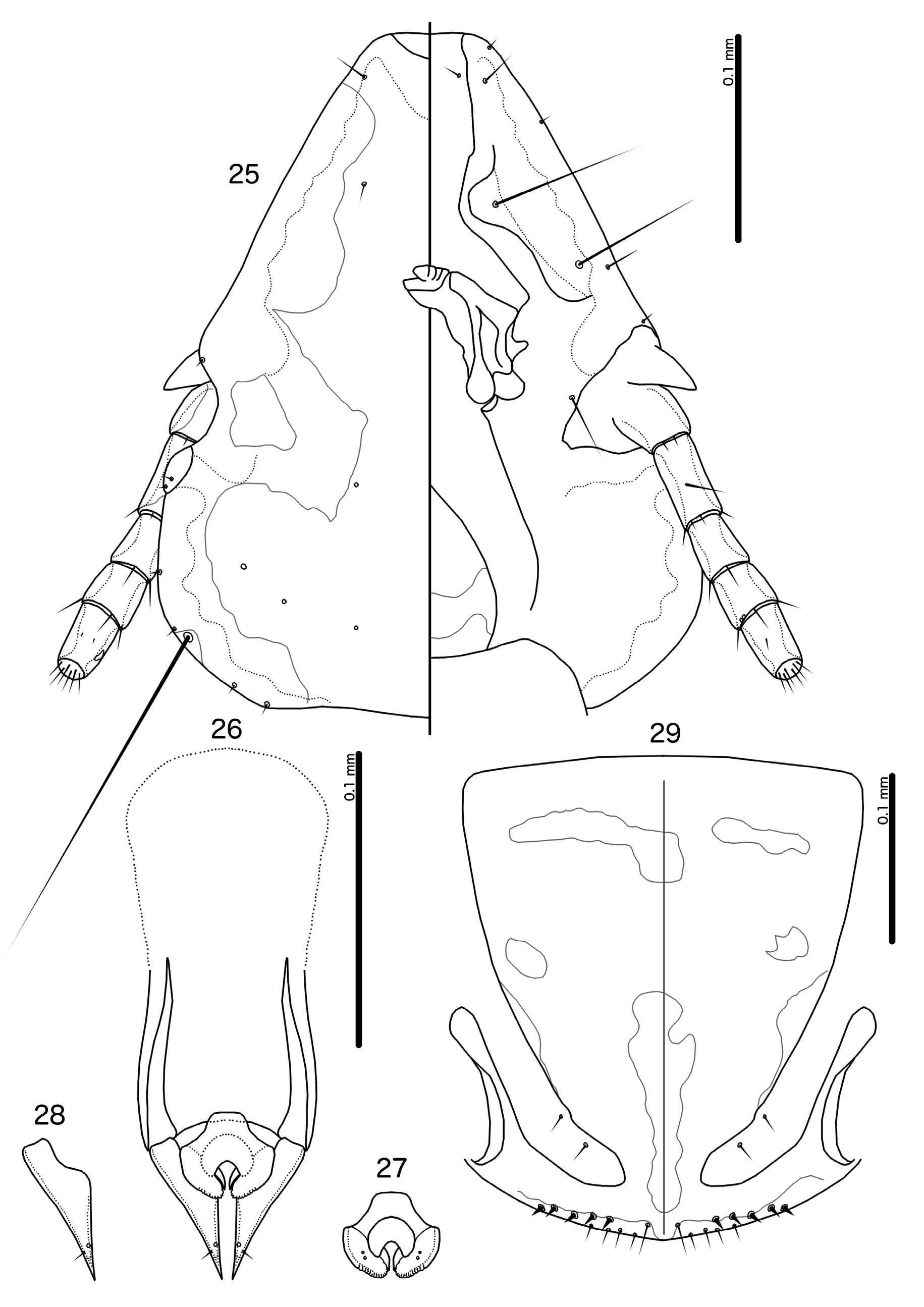
Figs 23–29 View Figs 23–24 View Figs 25–29 , 41–42 View Figs 37–44
Type host
Euplectes jacksoni (Sharpe, 1891) – Jackson’s widowbird ( Ploceidae ).
Type locality
Kenya.
Other host
Euplectes progne delamerei (Shelley, 1903) – long-tailed widowbird.
Diagnosis
Brueelia terpsichore sp. nov. belongs to the informal ʻAfrican pied Brueelia ʼ group (see above). Within this group, B. terpsichore sp. nov. is not particularly similar to any species. The extensive dark pigmentation of the sternites and subgenital plates of both sexes, the abdominal chaetotaxy, and the head shape suggests that B. aguilarae may be the closest relative of B. terpsichore sp. nov. These two species can be separated by the following characters: tps present on male tergopleurites VI–VIII in B. terpsichore sp. nov. ( Fig. 23 View Figs 23–24 ), but absent in B. aguilarae ; male abdominal segment IV with 2 ps on each side in B. terpsichore sp. nov. ( Fig. 23 View Figs 23–24 ), but with 1 ps on each side in B. aguilarae ; aps absent on male tergopleurite V in B. terpsichore sp. nov. ( Fig. 23 View Figs 23–24 ), but present in B. aguilarae ; proximal mesosome broadly rounded in B. aguilarae , but trapezoidal in B. terpsichore sp. nov. ( Fig. 27 View Figs 25–29 ); translucent fenestra of female subgenital plate T-shaped in B. aguilarae , but divided into smaller, isolated fenestrae in B. terpsichore sp. nov. ( Figs 29 View Figs 25–29 , 41 View Figs 37–44 ; note that there is some variation between specimens in this species); vulval margin gently rounded in B. terpsichore sp. nov. ( Fig. 29 View Figs 25–29 ), but convergent to median point in B. aguilarae .
Etymology
The specific epithet is in honor of the Greek muse Terpsichore, goddess of dance; ultimately from the Greek ʻ terpo ʼ for ʻI delightʼ and ʻ khoros ʼ for ʻdanceʼ. This refers to the peculiar lekking behaviour of the type host, which includes the construction of a small stage on which the male dances by jumping high into the air and singing to attract females ( Andersson 1989, 1991).
Material examined
Holotype KENYA • ♂, ex Euplectes jacksoni (as Drepanoplectes jacksoni ); Kenya; Jan. 1936; R. Meinertzhagen leg.; NHML 6084 (lower male on slide).
Paratypes KENYA • 2 ♂♂, 10 ♀♀; same data as for holotype; NHML 6081 View Materials , 6082 View Materials , 6084 View Materials (not lower male on slide), 6210 .
Other material KENYA • 1 ♂, 1 ♀, ex Euplectes progne delamerei (as Drepanoplectes progne delamerei ); Kenya; Jan. 1936; R. Meinertzhagen leg.; NHML 6714.
Description
Head rounded trapezoidal ( Fig. 25 View Figs 25–29 ), lateral margins of preantennal area slightly convex or almost straight, frons broadly flattened. Marginal carina broad, deeply displaced and much widened at osculum, with undulating median margin. Ventral anterior plate not visible. Head chaetotaxy and pigmentation pattern as in Fig. 25 View Figs 25–29 . Preantennal nodi large, bulging. Pre- and postocular nodi large. Marginal temporal carina broad, with distinctly undulating median margin. Gular plate slender, lanceolate. Thoracic and abdominal segments and pigmentation patterns as in Figs 23–24 View Figs 23–24 , 41–42 View Figs 37–44 .
Male
Thoracic and abdominal chaetotaxy as in Fig. 23 View Figs 23–24 . Anterior section of basal apodeme not pigmented and cannot be seen clearly in examined specimens; illustration here is approximate ( Fig. 26 View Figs 25–29 ). Proximal mesosome rounded trapezoidal, widening distally, with concave lateral margins ( Fig. 27 View Figs 25–29 ). Mesosomal lobes relatively slender, rounded distally, with rugose area limited to distal margin. Gonopore crescentshaped, slightly wider than long. Penile arms short, not reaching beyond distal margin of mesosome. Parameres elongated, tapering gently ( Fig. 28 View Figs 25–29 ); pst1–2 as in Fig. 28 View Figs 25–29 . Measurements ex Euplectes jacksoni (n = 3): TL = 1.42–1.48; HL = 0.33–0.34; HW = 0.26–0.28; PRW = 0.19; PTW = 0.26–0.27; AW = 0.36–0.37. Measurements ex E. progne delamerei (n = 1): TL = 1.44; HL = 0.34; HW = 0.26; PRW = 0.19; PTW = 0.27; AW = 0.35.
Female
Thoracic and abdominal chaetotaxy as in Fig. 24 View Figs 23–24 . Pigmentation pattern of subgenital plate differing slightly between specimens; two variants shown in Fig. 29 View Figs 25–29 (separated by grey line), one with anterolateral fenestra connected to antero-median fenestra and one with these fenestrae unconnected. Anteromedian fenestra generally pale brown in pigmentation, may extend posteriorly to approach posteromedian fenestra (not shown). Exact shape of postero-lateral fenestrae and translucent lateral borders of distal subgenital plate also differ between specimens. Subgenital plate rounded triangular, with moderate connection to cross piece. Vulval margin gently rounded, with 3–5 short, slender vms and 4–6 short, thorn-like vss on each side; 2–3 short, slender vos on each side of subgenital plate; distal 1 vos median to vss. Material from both host species with same vulval chaetotaxy. Measurements ex Euplectes jacksoni (n = 10): TL = 1.58–1.73 (1.66); HL = 0.36–0.38 (0.37); HW = 0.28–0.31 (0.30); PRW = 0.19–0.22 (0.21); PTW = 0.28–0.31 (0.30); AW = 0.40–0.44 (0.42). Measurements ex E. progne delamerei (n = 1): TL = 1.73; HL = 0.37; HW = 0.30; PRW = 0.21; PTW = 0.31; AW = 0.40.
Remarks
No significant differences have been found between specimens from the two host species, except that the single examined male from Euplectes progne delamerei lacks tps on tergopleurite VI. These are present in all specimens from the type host; however, the number varies between 1 and 2. As only one male from E. p. delamerei was examined, we presently do not attach any significance to this difference, as the psps on this segment is also missing on one side. The absence of these setae may thus be due to individual variation.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |