
Boroecia hopcrofti, Stępień & Błachowiak-Samołyk & Krawczuk & Angel, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4394.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:9358DA65-1111-4702-A898-52519D53B181 |
|
DOI |
https://doi.org/10.5281/zenodo.5958709 |
|
persistent identifier |
https://treatment.plazi.org/id/D723883B-FF93-FFCC-34BC-FF72FF5DBA4E |
|
treatment provided by |
Plazi |
|
scientific name |
Boroecia hopcrofti |
| status |
sp. nov. |
2.6. Boroecia hopcrofti new species
( Figures 16–18 View FIGURE 16 View FIGURE17 View FIGURE 18 )
Etymology. The species is named after Professor Russ Hopcroft, both in recognition of his considerable contributions to the study of zooplankton especially in the Arctic, and for facilitating our studies of the halocyprids from the Canadian Deep and from the Gulf of Alaska.
Type locality. Holotype: A male 2.64 mm long, dissected, stained with lignin pink and mounted on slides. Collected at Healy 0 502 station 13, 15th July 2005 at the type locality 75°15.8’N, 161° 12’W; depth 750– 500 m. Deposited at the Natural History Museum London NHMUK2016-628.
Paratype: A female 2.76 mm long, dissected, stained with lignin pink and mounted on slides. Collected in the same samples as the holotype. Museum registration number NHMUK 2016-629.
The other material examined is listed in Appendix 1.
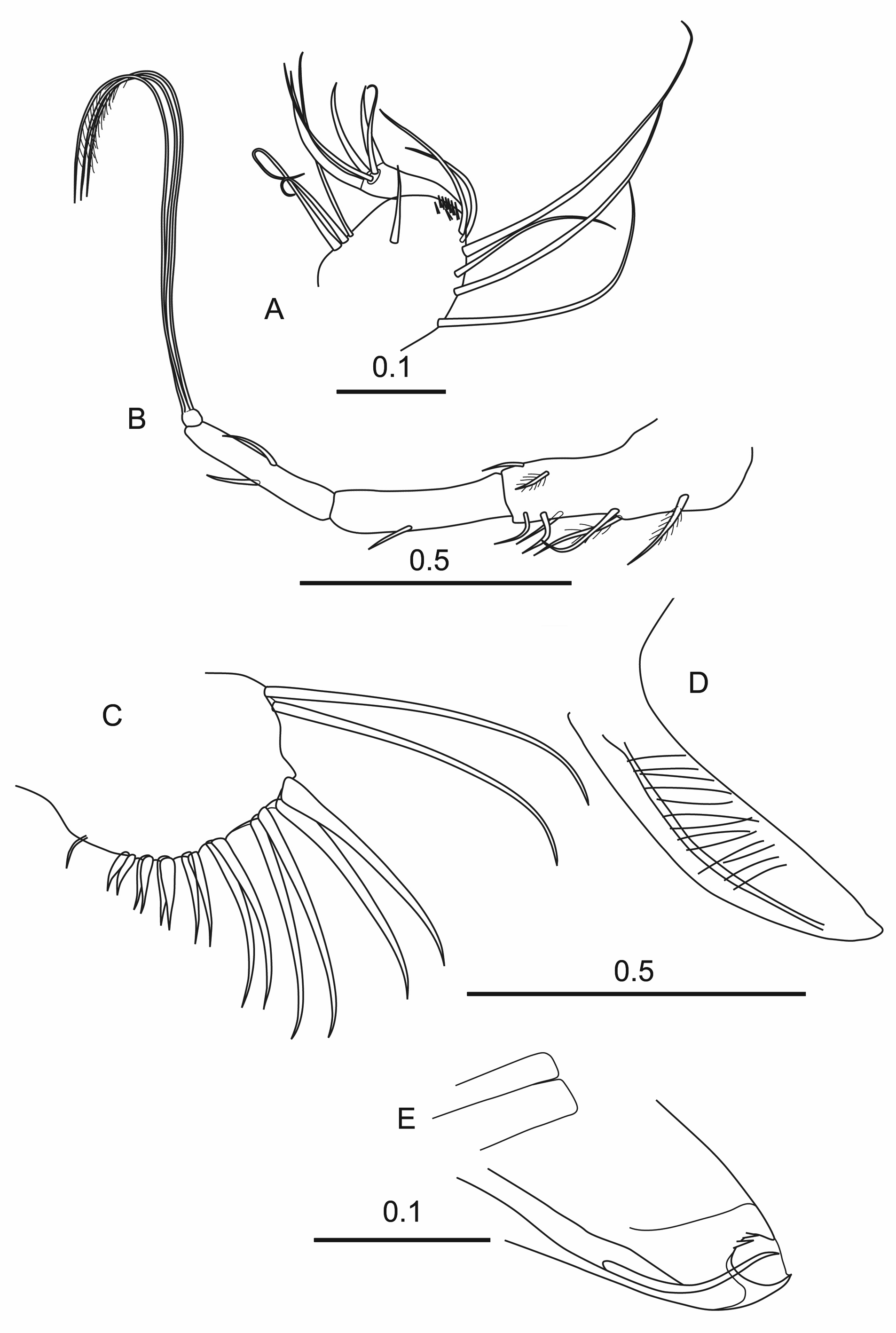
Male. Carapace ( Figures 16 A,B View FIGURE 16 ). The length is 2.67 ± 0.069 mm ( Table 1), the height, that is 40.0% CL is only slightly greater than the width (38.4% CL). The lateral outline is sub-rectangular with little or no anterior taper. The shoulder vaults are rounded and not strongly developed. In ventral view, the maximum width is just posterior to mid-length, and the anterior margin of the carapace below the incisure is slightly flanged. The rostrum, measured from the anterior end of the hinge between the valves, is 11–12% CL, which is slightly longer than the depth of the incisure (~8% CL). The left asymmetrical gland (LAG) opens just anterior to the posterior end of the carapace hinge (~4% CL) and twice that distance from the posterior margin of the carapace. The right asymmetrical gland (RAG) opens slightly above PVC (30–31% CL below the hinge-line). There is a group of lateral edge glands on both valves just dorsal to the RAG. The posterior dorsal corner of the carapace forms a rounded right angle on both valves, and lacks spines, although on the left valve there is a slight process that may be a residual spine. Beneath the incisure on the anterior flanks of the carapace are clear oblique striations; elsewhere there is no distinct ornamentation.
Frontal organ ( Figure 16 C View FIGURE 16 ). The stem (27.9% CL) just projects beyond the end of the limb of the first antenna. It is sutured just proximal to where the retaining setae lock the stem to the second segments of the A1; these setae are 9–10% CL behind the base of the capitulum. The capitulum is down-turned about 60°. It is elongate (13.4% CL) and spinose over most of its ventral margin, but only on the basal quarter of the dorsal margin. The end of the capitulum is bare and has a bluntly pointed tip
First antenna ( Figures 16 C,D View FIGURE 16 ). The segmentation is clear, and the basal segment is slightly longer than the second. Both segments are bare. The a-seta is convoluted and swollen at its base and carries a small appendage ( Figure 16 C View FIGURE 16 ). It is relatively long (18.5% CL) and extends back to mid-length of first segment. The b-seta (39.4% CL) has a short, ribbed section with 3–4 spines at its distal end. There is a further spine inserted close to the end of the seta. The c-seta (7.0% CL) is a little shorter than the second segment. The d-seta (31.5% CL) is quite short, barely extending beyond the end of e-seta armature. The e-seta (43.3% CL) has an armature of a double row, each with 34–35 broad hooked spines ( Figure 16 D View FIGURE 16 ), beyond this armature the seta is broadly flanged.
Second antenna ( Figures 16 E,F View FIGURE 16 ). Protopodite (45.3% CL) is nearly three times the length of the first exopodite segment (18.5% CL). In other Boroecia species there is a terminal seta that curls around the suture between the first and second segments of the exopodite, but this was not seen in the type specimen. Segments 2–9 of the exopodite, which carry the swimming setae, are about 40% the length of the first segment. The longest swimming seta is 43% CL and so slightly shorter than the protopodite. On the terminal segment there is a very short terminal seta (4.2% CL).
The first segment of the endopodite (7.3% CL) has quite a small processus mamillaris, which is slightly asymmetrical and bluntly pointed ( Figure 16 F View FIGURE 16 ). The a-seta is S-shaped, and shorter than the b-seta, which is almost straight and carries several long setules. The second endopodite segment is shorter than the first and the c- and dsetae have lengths similar to the width of the second endopodite segment. There is a thorn-like e-seta inserted at the base of f-seta. The f-seta (29.1% CL) is bare and distally slightly flanged. The g-seta (47.2% CL) is longer and more obviously flanged. The other h–j-setae (17–18% CL) are thin-walled, and the h-seta is inflated at its base. The hook appendage on the right limb has a long basal hasp, and angles back about 140°. The outer margin of the angle is slightly inflated. Beyond the angle, the hook is curved into a long arc reaching well beyond the base. The end of the hook is rounded and ribbed. In contrast, the hook on the left limb has a very short hasp and is right angled. Its distal part is almost straight and has a smooth end, tipped with small terminal thorn.
Labrum. The labrum has a shallow, curved depression and is flanked by about 22 filaments.
Mandible ( Figures 16 G,H View FIGURE 16 ). The first segment of the endopodite has a dorsal sub-terminal plumose seta, and ventrally one very long and three moderately long setae (i.e. not reaching to end of limb). The second segment has the three dorsal terminal setae (1 long, and 2 medium) characteristic for the subfamily, and ventrally two subterminal setae, one long and one medium. Terminally, the third segment has the usual seven setae, two of which are long curved spinose spines, the longest is relatively long with 19.3% CL. The toothed endite is also typical, it having two spine setae (one sharply pointed, the other rounded) and six subserrate triangular teeth, the first of which is offset from the others. There is also a very broad inner tooth that nearly completely overlaps three of the triangular teeth.
The inner face of the endite is lined with fine setules, and the exopodite is a short blunt process and carries a fairly short, densely plumose seta ( Figure 16 G View FIGURE 16 ). The coxale toothed edge consists of a broad tooth and nine other triangular teeth ( Figure 16 H View FIGURE 16 ). The outer tooth list is composed of two large curved spines followed by 16 small rounded teeth. The inner list consists of two large teeth, with three small teeth in-between, and a row of c.14 small blunt teeth.
Maxilla ( Figure 17 A View FIGURE17 ). The endopodite has two segments. The first has six setae on its anterior face, one lateral seta and three posterior setae; terminally on its outer surface are six elongate spines. The second segment has three terminal claw setae and close to them a pair of subequal tubular setae.
Fifth limb. There are three series of five setae on the epipodial appendage. The basale has the following setae: ventrally two groups of four setae (one plumose) and a group of two, laterally 1 + 1 setae, and dorsally a medium length plumose seta that extends beyond the midpoint of the first endopodite segment, and the long sub-terminal seta that is considered to be the remnant exopodite (Boxshall 1998), which extends to beyond the end of the limb. The first endopodite segment medially has two ventral setae and a dorsal seta all of which extend beyond the end of the segment. The terminal endopodite segment has three terminal claw setae with length ratios of 9:10:5, the longest central seta being 9.3% CL, and c. 60% the dorsal length of the limb.
Sixth limb ( Figure 17 B View FIGURE17 ). The epipodial setae consists of groups of 7 (one short) + 5 + 5 (ventral). The basale has six ventral setae (1 + 2 + 2 + 1), no lateral setae, and dorsally a short plumose seta and short sub-terminal seta. The first endopodite segment has a short medial seta on its ventral surface that does not extend to the end of the segment. Medially, the second segment has a single dorsal seta and a single ventral seta, which are also quite short. The third terminal segment carries three long, subequal, terminal setae all carrying long setules on the terminal quarter of their length.
Caudal furca ( Figure 17 C View FIGURE17 ). The furca carries the normal eight pairs of hook setae, with the first longest pair offset from the others. The first longest pair (18.5% CL) is relatively slim and curved throughout much of their lengths. They are armed with fine spines that are similar in length to the seta’s width. The next three pairs are stouter, and lined with thicker spines that are about a third width of the seta. The fifth pair is half the length of the fourth pair; this marked disparity in length between the fourth and fifth pairs of caudal furca spines is a characteristic of the genus. The last two pairs arch slightly forward and carry very fine spines.
Copulatory appendage ( Figure 17 D,E View FIGURE17 ). As is typical in halocyprids, this single structure is mounted on the righthand side of the caudal furca. Its anterior margin is almost straight, the posterior margin is strongly arched. The terminal part ( Figure 17E View FIGURE17 ) is curved and the tip is slightly hooked. Its length is 21.6% CL. There are 7 oblique muscles.
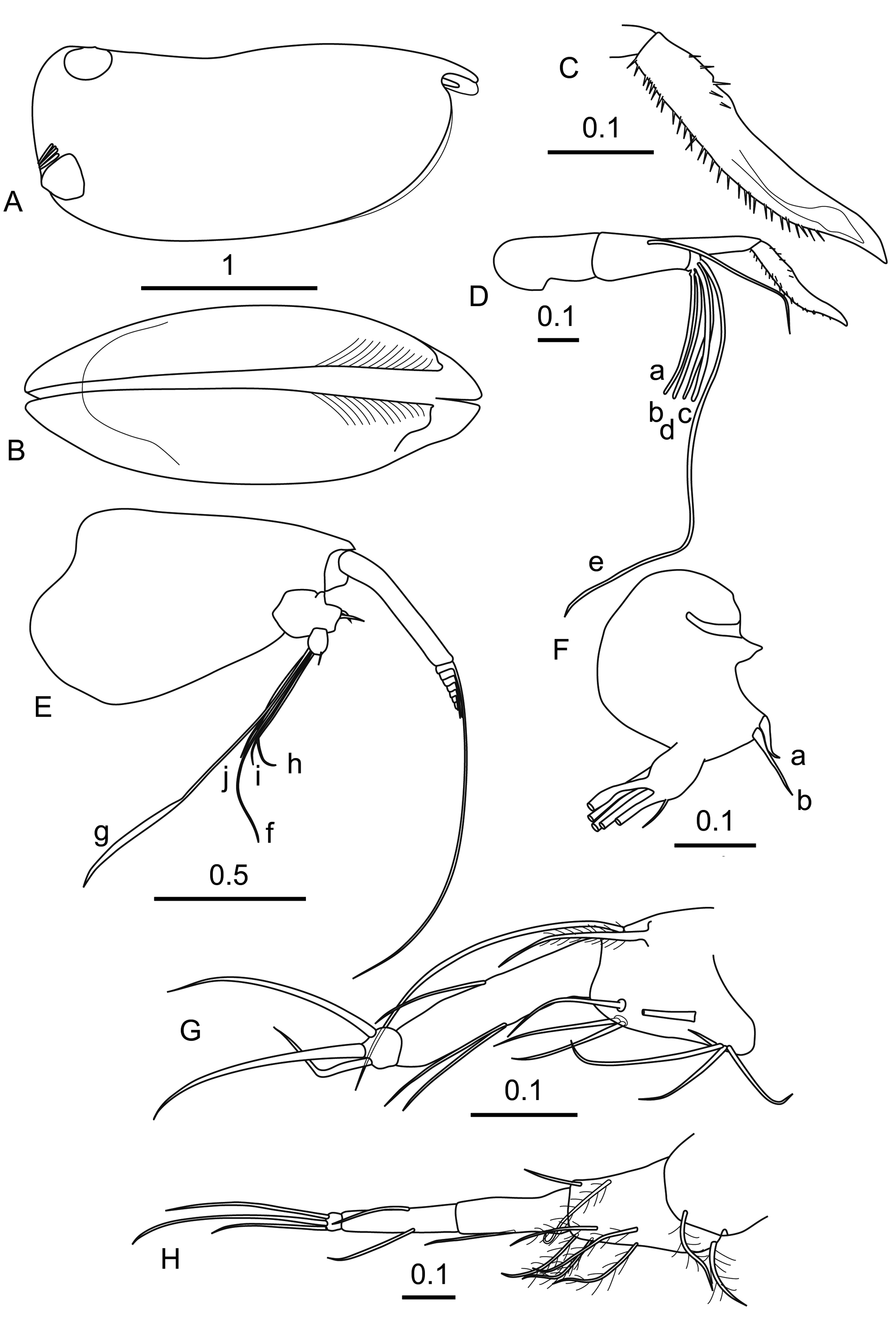
Female. Carapace ( Figures 18 A,B View FIGURE 18 ). The length is 2.89 ± 0.086 mm ( Table 1), the height is 45% CL and the width 40–41% CL. Viewed laterally, the female carapace is more tapered than the males with the maximum height at about two thirds length. The shoulder vaults are rounded and the posterior dorsal corner is a rounded right angle without spines. There is a clear pattern of oblique striae on the anterior part of the carapace below the incisure. The LAG opens just anterior (2.4% CL) to the dorsal end of the hinge between the carapace valves and about 7.5% CL anterior to the posterior margin. The RAG opens slightly above the PVC at about 85% the carapace height. Just above the RAG is a group of edge glands on both valves. The second group of edge glands, which are such a clear feature in B. maxima , are absent here). The length of the rostrum (11.7% CL) is greater than the depth of the incisure (8.3% CL).
Frontal organ ( Figures 18 C,D View FIGURE 18 ). The stem (19.2% CL) extends well beyond the end of first antenna. There is a clear suture between the capitulum and the stem. The capitulum ( Figure 18 C View FIGURE 18 ) (11.3% CL) is down-turned about 45° and S-shaped. It is spinose over most of its length ventrally, but only on the basal quarter of the dorsal surface. The tip is bare and quite sharply pointed.
First antenna ( Figure 18 D View FIGURE 18 ). The limb (18.8% CL) is only weakly segmented. There is a long dorsal seta (16.5% CL) on each of the second segments, reaching almost to the end of the capitulum. The a–d setae (10.9% CL) are 0.7 times as long as the e-seta. The e-seta (36.9% CL) is slightly flanged terminally and carries a sparse line of short fine spines along its trailing edge.
Second antenna ( Figure 18 E View FIGURE 18 ). The protopodite is 41.4% CL, and 2.5 times as long as the first exopodite segment. The first segment (16.2% CL) lacks the terminal seta seen in all other species in the genus. The remaining exopodite segments are 7.2% CL and carry the swimming setae, the longest of which is c. 43% CL. The first segment of the endopodite ( Figure 18 F View FIGURE 18 ) has a processus mamillaris, which is a blunt, curved cone. The a-seta is bare and curved near its tip. The longer b-seta is bare and nearly straight. The second endopodite segment has a cseta. The f-seta (21.8% CL) is slightly flanged. The g-seta (39.5% CL) is both, much longer and more clearly flanged. The h-, i- and j-setae are c. 17% CL.
Mandible, maxilla and labrum. All these limbs are similar to those of the male. The longest seta on the end of the mandible is 20% CL and nearly 80% the length of the endopodite.
Fifth limb ( Figure 18 G View FIGURE 18 ). On its ventral surface the basale has a proximal group of four (one plumose) setae proximally, and two more distally. There are two lateral setae, and dorsally a very long terminal seta that extends beyond the end of the limb and a long plumose seta that extends to two-thirds the length of the first segment. The first endopodite segment has the usual two ventral setae and one dorsal seta; all three extend to the end of the segment. The terminal segment has three setae. The dorsal seta is only slightly shorter than the medial seta, which is 7.6% CL and c. 48% the length of the limb. The ventral seta was broken.
Sixth limb ( Figure 18 H View FIGURE 18 ). The basale has three plumose ventral setae, two plumose lateral setae, and dorsally a plumose seta and a bare seta that is shorter than usual, reaching only to mid-length of the first segment of the endopodite. The first endopodite segment has only a single medial ventral seta. The second segment has single medial setae both dorsally and ventrally; both reach just beyond the end of the segment. The three terminal setae on the third segment are quite long and slim. Their length ratios (dorsal:central:ventral) are 8.5:10:5.5. The longest central seta is c. 14% CL, which is 60% of the length of the endopodite.
Caudal furca. Is similar to that of the male with the characteristic disparity between the fourth and fifth pairs of hook setae.
The carapace lengths of life history stages of B. hopcrofti are shown in Table 1 and compared with those of B. maxima from the same regions. Table 3 compares descriptive and meristic characters of the six described Boroecia species.
Distribution. B. hopcrofti was identified from all seven stations sampled in the Canadian Deep. In total 78 specimens of B. hopcrofti were identified. The number was far fewer than the 2359 specimens of B. maxima . Bathymetric range of B. hopcrofti tended to be a little deeper (300–500 m) than that of B. maxima (200–300 m). It was slightly more abundant at the slope stations (10 and 14) where the soundings were> 1000 m, but numbers were too small to be certain.
It has also been identified at just 4 stations collected at mesopelagic depths off the West coast of Svalbard ( Baczewska et al. 2012), and surprisingly, it was present in archived material collected in the Greenland Sea (see discussion). Hence it is probably far more widespread in cold waters of Arctic origin than these records might indicate.

| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Halocypridina |
|
Family |
|
|
SubFamily |
Conchoeciinae |
|
Genus |