
Lourinhasaurus, AS A
|
publication ID |
https://doi.org/10.1111/zoj.12113 |
|
persistent identifier |
https://treatment.plazi.org/id/A1521835-FFE1-FF8E-8334-61F35B83FC88 |
|
treatment provided by |
Marcus |
|
scientific name |
Lourinhasaurus |
| status |
|
LOURINHASAURUS AS A MEMBER OF MACRONARIA
Markedly opisthocoelus caudal dorsal vertebrae are synapomorphic for camarasauromorph sauropods ( Salgado et al., 1997; Upchurch et al., 2004) and are convergently shared by a few Chinese eusauropod taxa such as Mamenchisaurus ( Young, 1954; Young & Zhao, 1972; Ouyang & Ye, 2002) and Bellusaurus ( Dong, 1990) . The architecture of the middle dorsal neural spine of Lourinhasaurus is quite similar to that of some macronarians such as Camarasaurus ( Ostrom & McIntosh, 1966) . The distal transverse expansion of the middle and caudal dorsal neural spines observed in Lourinhasaurus and Camarasaurus was considered a synapomorphy of Macronaria by Wilson & Sereno (1998). Another feature recognized by D’Emic (2012) as a synapomorphy of the basal macronarian node Tehuelchesaurus +Titanosauriformes is the bifurcation of the pcdl in the middle and caudal dorsal vertebrae. This feature has been described in some dorsal vertebrae of Camarasaurus , Tastavinsaurus , and Cedarosaurus (Royo-Torres, 2009) , and also observed in Lourinhasaurus , suggesting a more widespread distribution among macronarians.
Lourinhasaurus bears a scapular blade with a large distal expansion, a feature that is also observed in basal macronarians such as Camarasaurus (e.g. Osborn & Mook, 1921; Ostrom & McIntosh, 1966) and Giraffatitan ( Janensch, 1961) . This feature differentiates Lourinhasaurus from Flagellicaudata members with an unexpanded distal scapular blade (e.g. Marsh, 1879, 1881; Osborn, 1904; Lull, 1919; Harris, 2007) and from the greater expansion of the genus Haplocanthosaurus ( Hatcher, 1903; McIntosh & Williams, 1988) and the distinct racquet-shaped distal expansion exhibited by rebbachisaurids ( Salgado et al., 2004; Whitlock, 2011).
A humerus with a squared proximal section, i.e. with a break of slope between its proximal and lateral margins that produces an acute or square angle proximolateral border, is variously considered in the literature. Wilson (2002) considered this character a synapomorphy of Somphospondyli (character 159 of Wilson, 2002), while D’Emic (2012) suggested that this feature is a synapomorphy of Macronaria (character 79 of D’Emic, 2012). However, basal macronarians such as Camarasaurus , Tehuelchesaurus , Brachiosaurus , and Lourinhasaurus are considered to present the plesiomorphic state, i.e. a rounded transition between the lateral and the proximal edges of the humerus. The co-planar distal ischiatic blade present in Lourinhasaurus is also considered a synapomorphy of Macronaria sensu Wilson & Sereno (1998). An unexpanded distal end of the ischium distinguishes it from that of diplodocoids such as Apatosaurus ( Marsh, 1879, 1881), Barosaurus ( Lull, 1919) , Dicraeosaurus ( Janensch, 1961) , and Diplodocus ( Hatcher, 1901) .
One of the forms closest to Lourinhasaurus is the macronarian Camarasaurus . This was noted earlier by McIntosh (1990a, b), and McIntosh et al. (1996b) suggested that the Moinho do Carmo sauropod was a putative new species of Camarasaurus , ‘ Camara- saurus ’ alenquerensis . Furthermore, some of the Camarasaurus autapomorphies ( Wilson & Sereno, 1998; Ikejiri, 2004) are shared by Lourinhasaurus . If the proposed close phylogenetic relationship between the two species is correct, these shared features have to be considered representative of a more inclusive group, Lourinhasaurus + Camarasaurus . Among the set of Camarasaurus autapomorphies shared by Lourinhasaurus are: (1) ischiatic blade directed caudally with its long axis passing through the pubic peduncle ( Wilson & Sereno, 1998); and (2) scapular bone with a rounded expansion on the acromial side ( Wilson, 2002). Ikejiri (2004) also provides a diagnosis for Camarasaurus , some of the features of which are shared by Lourinhasaurus , such as a massive pubis with short blade and a tibio-femoral length ratio of ∼0.66 (common in several sauropods, see D’Emic et al., 2013).
Camarasaurus is one of the best known sauropod genera; it comprises four species ( Upchurch et al., 2004): Camarasaurus grandis ( Marsh, 1877) , Camarasaurus lewisi ( Jensen, 1988) , Camarasaurus supremus Cope, 1877 and Camarasaurus lentus ( Marsh, 1889) . The abundant fossil record of Camarasaurus in the Upper Jurassic Morrison Formation has made it possible to identify the existence of great intraspecific variability related to ontogeny, sexual dimorphism, and individual variability ( Osborn & Mook, 1921; Ikejiri, 2004; Ikejiri, Tidwell & Trexller, 2005); such analysis has not been possible for other macronarians such as Tehuelchesaurus . It is necessary to compare Lourinhasaurus with the large range of variability shown by Camarasaurus to confirm its relationship with all members of the genus. In addition to the exclusive features supporting the validity of Lourinhasaurus and commented on above, there are some particular similarities and differences between the two taxa.
The cervical ventral surface of Camarasaurus is considered convex-to-flat transversely ( Upchurch et al., 2004, character 107; followed by other authors such as Carballido et al., 2011), which could suggest a different state than in Lourinhasaurus ; however, this codification is not sustained by the descriptions of several authors ( Osborn & Mook, 1921; Ikejiri, 2004) who describe a concave-to-flat ventral face for Camarasaurus , the same condition as is seen in Lourinhasaurus . In Tehuelchesaurus and Aragosaurus , it is not possible to verify this condition ( Sanz et al., 1987; Carballido et al., 2011); however, Europasaurus shares this condition with Lourinhasaurus and Camarasaurus ( Carballido & Sander, 2013) , and it is also present in Giraffatitan ( Janensch, 1950) and Galveosaurus ( Barco, 2009) .
Some features of the dorsal vertebrae, especially the neurapophyses system, are diagnostic for Camarasaurus ( Ikejiri, 2004, 2005). The vertebral centra of Camarasaurus and Lourinhasaurus show a general likeness, and the topology of pneumaticity is very similar (deep pleurocoels ramifying cranially and caudally). This type of bone tissue is also identified in Galveosaurus (considered as a macronarian form by Carballido et al., 2011) and Europasaurus ( Carballido & Sander, 2013) . In Europasaurus , the pleurocoel becomes less ramified along the vertebral series ( Carballido & Sander, 2013), and a similar pattern is observed in Lourinhasaurus , reducing the number of smaller cavities connected with the pleurocoels. Lourinhasaurus could be distinguished from Camarasaurus by the presence of a craniocaudal concavity on the ventral face of the cranial-to-middle dorsal centra; such a concavity is lacking in Tehuelchesaurus , Europasaurus , and Haplocanthosaurus ( Hatcher, 1903; Upchurch et al., 2004; Carballido et al., 2011; Carballido & Sander, 2013).
Several authors consider that the pcdl is not bifurcated or expanded in the caudal dorsal vertebrae of Camarasaurus (e.g. Upchurch et al., 2004). Nevertheless, the morphology of the pcdl in Camarasaurus is variable, and a bifurcated pcdl can be observed in the caudal dorsal vertebrae (e.g. Ostrom & McIntosh, 1966: pl. 25), as was indicated by Royo-Torres (2009). This derived condition is also present in other basal Macronaria such as Tehuelchesaurus ( Carballido et al., 2011) , Tastavinsaurus ( Canudo et al., 2008; Royo-Torres, 2009), and Cedarosaurus (Royo-Torres, 2009) but absent in Europasaurus ( Carballido & Sander, 2013) . The revision of the Camarasaurus laminae system will help in understanding its morphological variability in the time to access a codifica- tion in the data matrix. If the pcdl is really bifurcated in Camarasaurus , this feature could be a putative synapomorphy of the Camarasauromorpha node. The morphology of the preserved neural spine in Lourinhasaurus is similar to that of Camarasaurus (e.g. Osborn & Mook, 1921) and represents a unique form among sauropods. The circular depression in the cranial articulation is exclusive to Camarasaurus and Lourinhasaurus , but in Camarasaurus this central depression is neither constant nor circular.
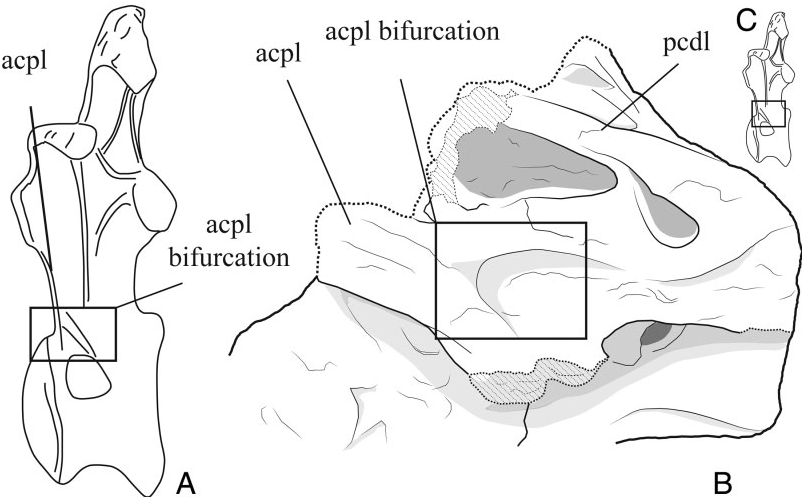
Camarasaurus and Lourinhasaurus share a particular feature: the presence of a bifurcation on the ventral tip of the acpl in which the caudal branch develops toward the cranioventral margin of the neural arch, bordering the pleurocoel dorsally ( Fig. 26 View Figure 26 ). This feature is also paralleled by other sauropods such as Tastavinsaurus (Royo-Torres, 2009, fig. 4.13; R. Royo-Torres, pers. observ., 2013). Other taxa such as Diplodocus and Dinheirosaurus exhibit a different condition in which the parapophyses are sustained by two laminae, a cranial one (acpl) and a caudal one (pcpl) ( Osborn, 1899; Hatcher, 1901; McIntosh & Williams, 1988; Bonaparte & Mateus, 1999; Mannion et al., 2012). The presence of pcpl in dorsal vertebrae is recognized in several neosauropods ( Wilson, 2002) and in some eusauropods such as Jobaria ( Sereno et al., 1999) and turiasaurs ( Royo-Torres et al., 2006). The pcpl is absent in Camarasaurus and in the preserved dorsal neural arches remains of Lourinhasaurus .
In the proximal caudal vertebrae, similarities are also observed in the morphology of the centra and the neurapophyses. These vertebrae share the main centra morphology, the triangular transverse process, and the neural spine morphology. Lourinhasaurus could be distinguished by the presence of a smooth concave ventral face and a less constricted centrum; nevertheless, the interpretation of the caudal vertebrae ventral face is problematic. In character 183 of Upchurch et al. (2004) data matrix, the ventral face of Camarasaurus proximal caudal vertebrae was codified as transversely convex. By contrast, Ikejiri (2005) described this ventral face as flat, which is similar to the Lourinhasaurus state. Europasaurus also presents flat to slightly concave ventral faces on the most proximal caudal vertebrae ( Carballido & Sander, 2013). The slight ventral concavity present in Lourinhasaurus caudals does not resemble the marked concavity common in diplodocoids and in some titanosaurs ( Upchurch et al., 2004). This situation suggests that it may be necessity to revise the character definition, considering the presence of at least three different states: (1) a clear convex ventral face (the presence of a craniocaudal hollow); (2) a marked concave ventral face; (3) and a flat ventral face (incorporating slight convex and concave states – this state could be identified by a break in the slope between the lateral and ventral faces of the centrum).
The proximal caudal neurapophyses are transversely compressed in both taxa. However, the Lourinhasaurus neurapophysis differs from that of Camarasaurus by the presence of a circular and deep sprf. The fan-like morphology of the neural spines, in cranial view, is solely shared with C. grandis and C. supremus ( Ikejiri, 2004; Ikejiri et al., 2005). Wilson (2002, character 123) accepts the presence of prespinal laminae in Camarasaurus proximal caudal spines. Nevertheless, it is herein considered that the cranial surface of the Camarasaurus caudal spines bears a shallow and wide prespinal process that is particularly different from the stout and well-defined prespinal lamina present in diplodocids and several titanosauriforms ( Salgado et al., 1997; Wilson, 1999; Wilson et al., 2011). Lourinhasaurus and other sauropods such as Aragosaurus ( Sanz et al., 1987) , Spinophorosaurus ( Remes et al., 2009) , and Tastavinsaurus ( Canudo et al., 2008; Royo-Torres, 2009) also share the presence of a similar wide prespinal process. Wilson (2002, character 124) also refers the presence of a postspinal lamina in the caudal face of the proximal caudal spines of Camarasaurus , but the available figures do not confirm the presence of this lamina. Lourinhasaurus lacks a postspinal lamina in proximal caudal neural spines.
The subrectangular shape of the scapular distal portion in Lourinhasaurus is markedly distinct in most sauropods and is shared only by Camarasaurus grandis ( Ostrom & McIntosh, 1966) and a specimen of C. supremus (BS-179, Ikejiri, 2004). Ikejiri (2004) also noted great intraspecific variability in scapulae of Camarasaurus members. The articulation between coracoids and scapulae bears an indentation in Lourinhasaurus that does not occur in Camarasaurus ( Ostrom & McIntosh, 1966; McIntosh et al., 1996a). This feature is also observed in Giraffatitan ( Janensch, 1961) and Spinophorosaurus ( Remes et al., 2009) . The Lourinhasaurus humeri can be distinguished from the humeri of C. supremus ( Osborn & Mook, 1921) and C. lentus ( Ikejiri, 2004) by its more asymmetrical proximal portion (with medially pointed humeral head) and by a lesser value for mediolateral diaphysis/mediolateral proximal portion length. By contrast, the Lourinhasaurus humerus shares these conditions with C. grandis (e.g. Ostrom & McIntosh, 1966; McIntosh et al., 1996b) and C. lewisi ( McIntosh et al., 1996b) . McIntosh (1990a, b) suggested the necessity of establishing a new genus based on humerus/femur length ratio, which is higher in Lourinhasaurus than in Camarasaurus . In fact, the data provided by Ikejiri (2004) show that there is a sub-adult of Camarasaurus ( C. lentus, USNM 13786) with a high humerus/femur length ratio.
The ulna is another distinct bone compared with that of Camarasaurus . The Lourinhasaurus ulna shares some features with Titanosauriformes ( D’Emic, 2012) that are absent in Camarasaurus and Tehuelchesaurus . One of these features is a caudally unexpanded distal end present in Lourinhasaurus and Titanosauriformes. Camarasaurus has a marked caudal expansion of the distal ulnar end ( Ostrom & McIntosh, 1966) that is shared by several non-Titanosauriformes sauropods ( D’Emic, 2012). In proximal view, Lourinhasaurus also shows a craniomedial process that is longer and more robust than the craniolateral process. The carpus II of Lourinhasaurus is comparably higher than the carpus II of Camarasaurus ( Osborn, 1904; Ostrom & McIntosh, 1966).
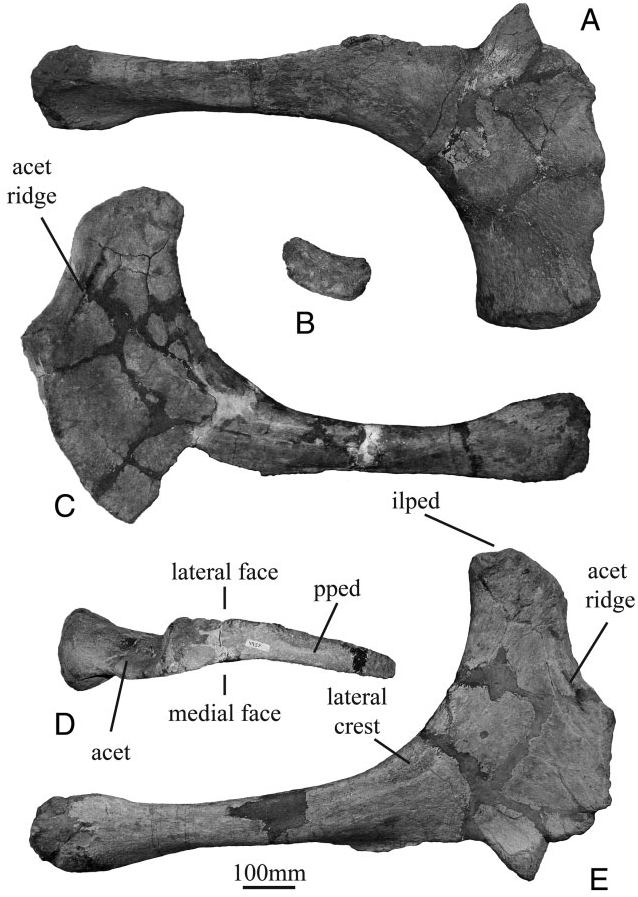
The ilium exhibits some features that are interpreted as distinct from Camarasaurus , such as a caudally orientated postacetabular process, a ridge on the ventral margin of the postacetabular process and a small ridge along the ventral margin of the preacetabular process near the pubic peduncle. Lourinhasaurus exclusively shares a pronounced caudal deflection of the ischiatic blade with Camarasaurus , most precisely with Camarasaurus grandis ( Ostrom & McIntosh, 1966) ( Fig. 20 View Figure 20 ) and C. lentus (YPM 1910, McIntoch, 1990b). This caudal deflection is more pronounced than is found in Aragosaurus ( Sanz et al., 1987) , Camarasaurus lewisi ( McIntosh et al., 1996b) , C. supremus ( Osborn & Mook, 1921) , C. lentus (WDC A-BS-9 and WDC BS-615, Ikejiri, 2005), and Tehuelchesaurus ( Carballido et al., 2011) . Another particular feature of the ischium is the presence of a lip in the pubic articulation, a feature that is also shared by a specimen of Camarasaurus supremus ( Osborn & Mook, 1921: fig. 94). Nevertheless, the absence of this feature in other C. supremus ischia suggests the presence of intraspecific variability for this character.
Recent phylogenetic studies are recovering a few classical and new sauropod specimens as basal Macronaria ( Sander et al., 2006; Carballido et al., 2011; Whitlock, 2011; D’Emic, 2012; Carballido & Sander, 2013; Mannion et al., 2013). Haplocanthosaurus, which is considered by some phylogenetic hypotheses to be a basal macronarian ( Wilson & Sereno, 1998; Upchurch et al., 2004; Royo-Torres et al., 2006; Wilson & Upchurch, 2009; Carballido et al., 2011; Carballido & Sander, 2013), can be easily distinguished from Lourinhasaurus by the presence of a marked distal expansion of the scapular blade, absence of the opisthocoelous condition up to the sacrals and the presence of pcpl on the dorsal vertebrae ( Hatcher, 1903; McIntosh & Williams, 1988). Some recent studies (e.g. Remes et al., 2009; Whitlock, 2011) recovered Haplocanthosaurus as member of Diplodocoidea.
Europasaurus has recently been considered a basal macronarian in several analyses (e.g. Carballido & Sander, 2013) and is dated to the middle Kimmeridgian, slightly older than Lourinhasaurus (late Kimmeridgian to early Tithonian). In addition to its small size, in which it differs from other basal macronarians, it presents distinct aspects compared with Lourinhasaurus . One of the most marked differences between the two taxa is morphology of the dorsal neural spines, which have less transversely expanded distal tips and developed pre- and postspinal laminae. Europasaurus proximal caudal vertebrae bear a concave caudal articulation ( Carballido & Sander, 2013), thus differing from other macronarians with flat-to-concave caudal surfaces such as Camarasaurus ( Osborn & Mook, 1921) , Lourinhasaurus , Brachiosaurus ( D’Emic, 2012) , Haplocanthosaurus ( D’Emic, 2012), Cedarosaurus ( Tidwell et al., 1999) , and Aragosaurus ( Sanz et al., 1987) . Tastavinsaurus has a flat surface that bears a central concavity (Royo- Torres, 2009) as in Lourinhasaurus . Europasaurus also presents distinct features from Lourinhasaurus in its appendicular skeleton. One of the most important differences occurs in the ilium, where Europasaurus presents a developed subcircular preacetabular process that is common in the titanosauriformes ( Wilson, 2002).
Tehuelchesaurus is a basal macronarian from the Cañadón Calcáreo Formation ( Carballido et al., 2011), dated as Oxfordian–Kimmeridgian ( Cúneo et al., 2013). The phylogenetic hypotheses proposed herein suggest a close relationship with Camarasaurus and Lourinhasaurus , making both taxa a monophyletic clade, Camarasauridae . Nevertheless, Tehuelchesaurus can be distinguished in several ways from Lourinhasaurus and Camarasaurus . Tehuelchesaurus can be differentiated from Camarasaurus by the absence of a ventrally unbifurcated spol and from Lourinhasaurus and Camarasaurus in several aspects: (1) lack of distal expansion of the scapular blade, as in the flagellicaudatans (e.g. Marsh, 1879; Ostrom & McIntosh, 1966; McIntosh, 2005; Remes, 2006; Harris, 2007); (2) robust ulna and radius; (3) transversely compressed pubic peduncle; (4) the absence of pronounced ischiatic blade caudal deflection; and (5) femur with subcircular condyles and a slight lateral bulge.
Among the species of Camarasaurus , there is great morphological disparity. Camarasaurus grandis shares the highest level of similarity with Lourinhasaurus alenquerensis . In further analysis, the incorporation and recodification of every Camarasaurus species will be necessary to test the relationship of Lourinhasaurus with the described species of Camarasaurus and to confirm the validity of Lourinhasaurus as a valid genus or as a species of Camarasaurus , and then to evaluate the implications for Camarasaurus palaeobiogeography.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Lourinhasaurus
| Mocho, Pedro, Royo-Torres, Rafael & Ortega, Francisco 2014 |
Dinheirosaurus
| Bonaparte & Mateus 1999 |
Camarasaurus supremus
| Cope 1877 |