
Bairdoppilata hirsutella Maddocks, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5175.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:44FB9C3D-3188-4BFB-BDB8-C1324729A396 |
|
DOI |
https://doi.org/10.5281/zenodo.7008022 |
|
persistent identifier |
https://treatment.plazi.org/id/03FE6B50-FFE5-FFBE-ECD6-ADDB6F231A85 |
|
treatment provided by |
Plazi |
|
scientific name |
Bairdoppilata hirsutella Maddocks |
| status |
sp. nov. |
Bairdoppilata hirsutella Maddocks View in CoL , n. sp.
( Figures 11 View FIGURE 11 , 12A–M View FIGURE 12 , 13A–F View FIGURE 13 , 14A View FIGURE 14 )
? 1969 Bairdoppilata ( Bairdoppilata ?) hirsuta (Brady) .—Maddocks, p. 79, fig. 43A–I, pl. 2, figs. 1, 2 [specimens USNM 121353 and 121355 from the Gulf of Mexico, only, with uncertainty].
1973 Bairdoppilata hirsuta (Brady) .—Maddocks, p. 42, figs. 5B–G, 6A–E.
Material Examined: Eleven specimens, including USNM 139893, 139895–139900 ( Maddocks 1973).
Types: Holotype male specimen 634M, USNM 139891 About USNM ; illustrated paratype female USNM 635 About USNM F, USNM 139892 About USNM ; illustrated paratype, molting juvenile specimen 689J, USNM 139894 About USNM .
Type locality: Eltanin Station 25 in the Pacific Ocean, east of the Galapagos Islands ; 04 o 53’N, 80 o 28’W, depth 2489 m.
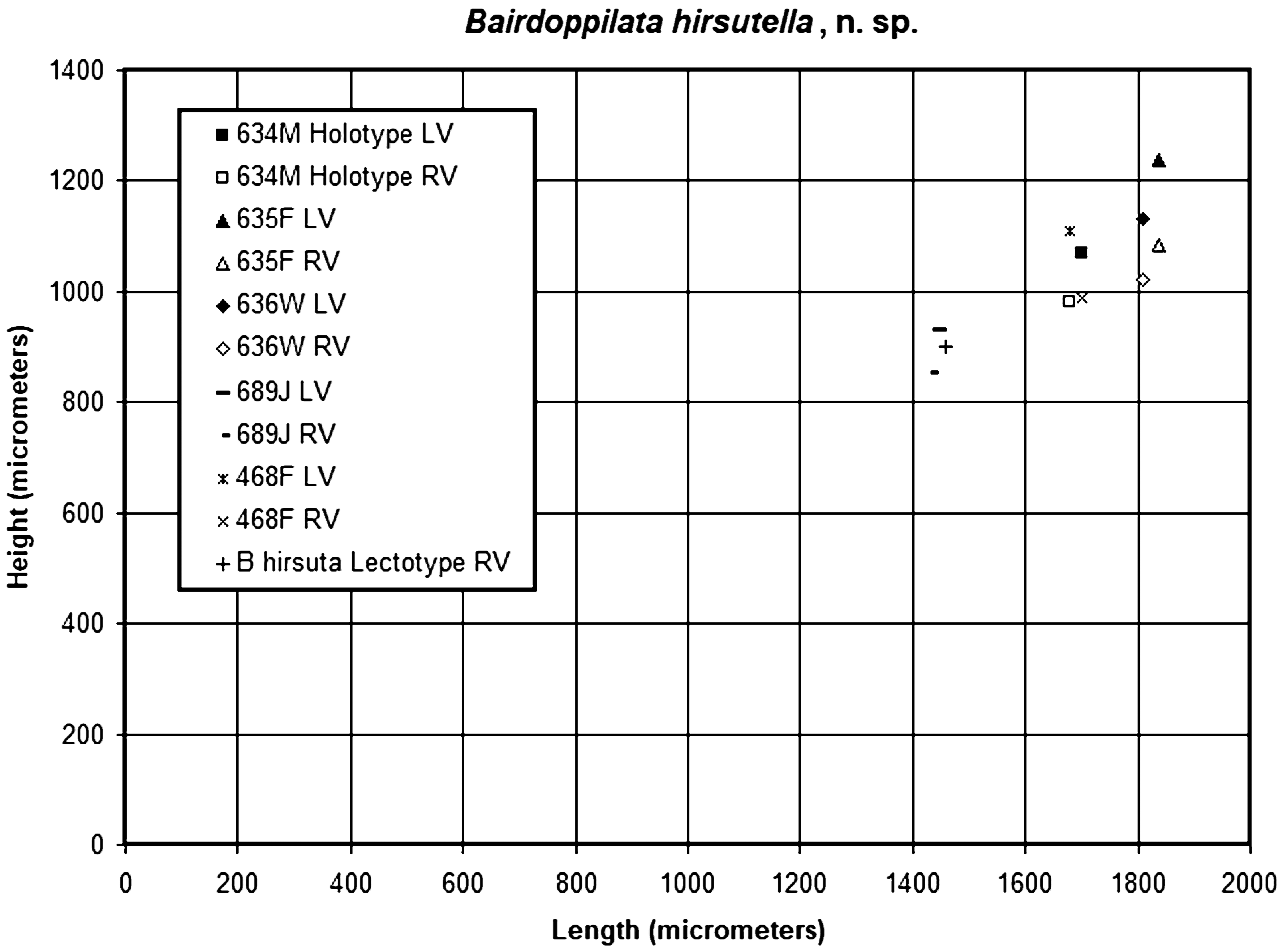
Dimensions: Male specimen 634M (USNM 139891): LVL 1.700 mm, LVH 1.070 mm, RVL 1.680 mm, RVH 0.980 mm. Female specimen 635F: USNM 139892: LVL 1.836, LVH 1.238, RVL 1.836 mm, RVH 1.083 mm. Molting juvenile specimen 689J: USNM 139894: LVL 1.447 mm, LVH 0.930 mm, RVL 1.434 mm, RVH 0.853 mm. See also Fig. 11 View FIGURE 11 .
The RV dimensions of juvenile specimen 689J are close to those reported for the RV lectotype of B. hirsuta by Puri & Hulings (1976), but the latter was described as an adult .
Esophageal Valve: The plate ( Fig. 13C View FIGURE 13 ) is broad, thin, and gently curved with smooth contours. There are about 18 low, thin, conical to subcylindrical teeth, which are widely spaced around the perimeter; the middle teeth are the smallest, and sizes increase somewhat toward the corners. There are no corner teeth, and the corners are smoothly rounded.
Anatomical Remarks: The rotund carapace is inflated, thickest at mid-height near the AMS, and smooth, glossy, with NPC but no surface sculpture ( Figs. 12H–M View FIGURE 12 ). In lateral outline it is broadly oval and upright in posture, with evenly curved dorsal and ventral margins. Both the anterodorsal corner and the caudal process are located distinctly above mid-height. Bairdoppilatan dentition is well developed ( Figs. 12F–G View FIGURE 12 ). The calcified inner lamella of the A-1 instar is fairly broad ( Figs. 12A, C View FIGURE 12 ) but weakly calcified and easily damaged.
The adult A2 and walking legs are extraordinarily long and thin, as usual in deep-sea animals. The distal claws of the A2 ( Fig. 13D View FIGURE 13 ) are equal in size, long and very thin. The fused claw is perfectly smooth and tapers to a sharp point.
Maddocks (1973, fig. 5B–G, 6A–E) published a full suite of drawings for male specimen 634M, USNM 139891 About USNM , which is here designated as the holotype. The drawing of the hemipenis showed the non-erect condition, with several overlapping processes enveloped by the hood-like distal appendage. Photographs of the same specimen are provided here as Figs. 13A–D View FIGURE 13 .
The conspicuous carapace sensilla ( Fig. 12H View FIGURE 12 ) for which B. hirsuta was named are not unique to this species or genus but are seen in many bairdioids (for example, fig. 19N of Brandão, 2008). They are more noticeable in deepsea forms, because the dark color contrasts with the stark white valves. The number of NPC increases for each instar in bairdiids ( Smith & Kamiya 2002). At each molt, the existing sensilla are retained, although their exocuticular covering is discarded with the exuvia. The inherited sensilla are thicker, longer, and darker in color than those newly added. On the adult carapace there are numerous NPC of different sizes, bearing sensilla of several ontogenetic ages, as indicated by length, thickness and color. One may speculate that this canopy of sensilla allows a small animal to claim a large volume of space, confers buoyancy, provides early warning of predation, and protects the carapace from abrasion and siltation.
Taxonomic Remarks: Maddocks (1973) identified these specimens as B. hirsuta (Brady) , but that identification requires re-examination. The soft parts of B. hirsuta are unknown, the lectotype may be a juvenile, the type locality is many thousands of kilometers distant, and no geographically intermediate populations have been reported. Brandão (2008) demonstrated that the diversity of deep-sea bairdioids has been greatly under-estimated, and it is best to be more conservative with identifications of Brady’s species. For clarity of communication, the population collected at Eltanin station 25 is named here as a new species.
B. hirsutella is similar to the two females collected from the Gulf of Mexico, which were identified as B. (B.?) hirsuta by Maddocks (1969) (specimens 468F, 469F, USNM 121353 About USNM , 121355 About USNM ). Their identity is considered uncertain at present. Larger assemblages, including males, will be required to evaluate the relationship of these distant populations .



| USNM |
Smithsonian Institution, National Museum of Natural History |
| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Bairdioidea |
|
Family |
|
|
Genus |