
Bachia geralista, Jr, Mauro Teixeira, Recoder, Renato Sousa, Camacho, Agustín, Sena, Marco Aurélio De, Navas, Carlos Arturo & Rodrigues, Miguel Trefaut, 2013
|
publication ID |
https://doi.org/10.11646/zootaxa.3616.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:D6F85F14-2D4E-4B53-9B40-3B6E9BE723FF |
|
DOI |
https://doi.org/10.5281/zenodo.5627975 |
|
persistent identifier |
https://treatment.plazi.org/id/0384FD70-5067-3268-26A4-7E18FEF5BD09 |
|
treatment provided by |
Plazi |
|
scientific name |
Bachia geralista |
| status |
sp. nov. |
Bachia geralista sp. nov.
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Bachia bresslaui Rodrigues et al. (2008:45)
Holotype: MZUSP 99408, an adult from Parque Nacional das Cavernas do Peruaçu, ( 15°09'14.04"S, 44°18'4.32" W, 740 m a.s.l., WGS84), Januária municipality, state of Minas Gerais, Brazil, collected by the authors on 25th January, 2008. Field number MTJ 212.
Paratopotypes: MZUSP 99406, 99407, 99409, 103211–10321, all collected by the authors, in January and July 2008, and January and July 2009.
Paratypes: MZUSP 100021, an adult from Roda Velha ( 12°47'1.86"S, 45°56'12.77"W, 750 m a.s.l.), São Desidério municipality, state of Bahia, Brazil, collected by M.A. Freitas on 5th January, 2010. MZUSP 94473–94474, Parque Nacional Grande Sertão Veredas ( 15°13'0.12"S, 45°47'60.00"W, 670 m a.s.l.), Formosa municipality, Minas Gerais, Brazil, collected by C. Nogueira on 23–29th October, 2001.
Etymology: The specific name is a noun in apposition; “ geralista ” is an old Brazilian term applied to the inhabitants of the “Planalto dos Gerais” from which the new species seems to be endemic.
Diagnosis: (1) A large-sized species of Bachia ( 101 mm of maximum known SVL); (2) prefrontals absent; (3) six supralabials; (3) 2-2 supraoculars and 2-2 supraciliars; (4) 2-2 femoral and 1-1 preanal pores; (5) 33–35 scales around midbody; (6) 46–51 dorsal rows of scales; (7) 37–40 ventral rows of scales; (8) 8–9 gulars; (9) 5 large preanal scales, and two small, on each side of the preanal plate; (10) sixth supralabial in contact with parietal, in most of the individuals; (11) ventral scales quadrangular, smooth; (12) fore and hindlimbs ending in an single apical scale; (13) first temporal absent.
Description of the holotype: Body elongate, with a slight cervical constriction on head, snout highly prominent and wedge shaped, tail longer than body. Rostral broad, prominent, contacting first supralabial, nasal and frontonasal. Viewed from above the rostral is about twice as wide as high; on lateral view it projects broadly anteriorly toward, forming a horizontal surface ventrally, at the level of ventral surface of the upper lips. Frontonasal trapezoidal, as wide as long, wider posteriorly, contacting rostral, nasal, first supraocular and frontal. Prefrontals absent. Frontal pentagonal, longer than wide, with anterior margin slightly convex, narrow than, and in broad contact, with frontonasal; lateral margins straight to slightly concave, in contact with first supraocular; posteriorly angulose, broadly contacting parietals and in short contact with interparietal. Frontal about three times wider than anterior supraocular. Frontoparietals absent. Interparietal narrow, longer than wide, subrectangular, slightly wider posteriorly, shorter than parietals, as long as frontal. Parietals very large, longer than wide, longer and wider than frontal, roughly hexagonal; their anterior margin deeply indented and in broad contact with frontal, contacting narrowly the first and broadly the second supraoculars, the postocular, punctually the sixth supralabial, two temporal and the dorsals; internally it contacts frontal and interparietal. Posterior borders of interparietal and parietals and dorsals coincides with a very slight transverse cervical constriction in the occipital region. Two supraoculars, first largest, about three times longer than wide, contacting frontal, frontonasal, nasal, loreal, first superciliary, second supraocular and parietal. Second supraocular smaller, above second superciliary, contacting postocular, longer than wide, separated from frontal by the slight contact between parietal and first supraocular. Two superciliaries, the first longer, their suture coincides with that between supraoculars. Nasal large, longer than high. Nostril in the middle of lower margin of nasal. Loreal roughly squared, in contact with nasal, first supraocular, first superciliary, preocular and second and third supralabials. Preocular small, curved. Frenocular present in contact with subocular. Six supralabials, third, fourth and fifth under the orbital region, sixth the highest and largest, contacting punctually parietal. One long subocular. Eyelid present with an undivided semitransparent disc. A large and elongate postocular between fifth and sixth supralabials and parietal. First temporal absent. Second temporal enlarged, longer than wide, temporal scale between parietal and sixth labial, in broad contact with both. Ear opening absent. All head scales smooth and juxtaposed.
Mental roughly trapezoidal, wider than long, wider than the ventral surface of rostral. Postmental heptagonal, as wide as long. Two pairs of chin shields, both contacting infralabials; the anterior pair smaller, in broad contact at midline; second pair in narrower contact at the midline; a pair of symmetric flat and diagonally disposed elongate pregulars. Five infralabials, first the smallest, second, third and fourth with about the same size. Gulars smooth, imbricate, rounded posteriorly, in nine transversal rows; scales of gular rows increasing gradually in size toward interbrachial region. Interbrachial region with four scales, the central ones largest, twice longer than wide. Lateral scales of neck subrectangular, smooth, imbricate, slightly rounded posteriorly and longer than wide, disposed in regular transverse rows and becoming gradually similar to adjacent dorsal or ventral scales. Collar fold absent.
Dorsal scales imbricate and disposed in regular transversal rows; smooth, subrectangular and wider in occipital region, becoming progressively narrower, more elongate and rounded towards the level of the forelimbs and then on longer, hexagonal, lanceolate, strongly keeled, with lateral sides almost juxtaposed. Fifty transverse rows between interparietal and the level of hind limbs. Lateral scales about the same size as dorsals but smooth and less acuminate; those closer to ventrals slightly wider. A distinctive area with granular scales surrounds the area of arm insertion and the posterior part of leg insertion. Thirty two scales around midbody. Ventral scales smooth, longitudinally imbricate, laterally juxtaposed, almost squared just after the interbrachial row, becoming gradually longer than wide, rounded posteriorly, those after midbody narrower; 38 transverse rows between interbrachials and preanals. Seven scales (five larger, central scales, and one small placed in each side of the preanal plate) in the posterior part of preanal plate, central one largest. One preanal and two femoral pores on each side.
Scales of tail similar to midbody dorsals, keeled, lanceolate, strongly imbricate.
Fore limbs rudimentary, stiliform, covered by smooth and imbricate scales, ending in a single apical scale. Its length corresponds to one row of lateral scales width. Hind limb also stiliform but larger than fore limb, covered by smooth, large and imbricate scales ending by one apical scale; its length equivalent to one and half scale rows. Two femoral pores present at each side, proximal one the largest.
Background dorsal and lateral surfaces of body and tail cream to light brown, with a dorsolateral light stripe. Ventral parts of body and tail cream immaculate.
Measurements of the holotype (mm): SVL=87.86; IBNL=15.91; LBM=67.90; HH=4.08; HW=5.31; HL=8.90, FLL=2.44; HLL=3.25; END =3.96, NND=2.60; TL=140 (regenerated)
Variation: Most individuals show the same color pattern as described for the holotype, however larger individuals tends to have a more homogeneous coloration, with faded stripes ( Fig. 2 View FIGURE 2 A,B). Although none individual possess a first temporal (Figs. 1,3), in some specimens none supralabial is in contact with parietal, because the postocular is in contact with the second temporal row. In some individuals in one side of the head there is contact between supralabial and parietal and in the other side there is not. There is also variation in the number of scales on the second temporal row: in some individuals the two scales on the second temporal row are merged. Meristics and morphometric values of B. geralista sp. nov. and B. bresslaui are given on Table 1 View TABLE 1 .
Meristics Morphometrics
B. geralista sp. nov B. bresslaui B. geralista sp. nov B. bresslaui *Only smaller adult individuals had their tails intact, thus this range may not be complete.
Comparison with other species (data from species in comparison are given in parentheses): the new species is a member of the B. bresslaui group by having lanceolated, keeled, imbricate dorsal scales, quadrangular, juxtaposed ventrals, 2-2 femoral pores, 1-1 preanal pores, limbs ending in one apical scale. These characters promptly distinguish B. geralista sp. nov. from all species of the B. dorbignyi group ( B. dorbignyi (Dumeril & Bibron, 1839) B. barbouri Burt & Burt, 1931 , B. bicolor (Cope, 1896) , B. huallagana Dixon, 1973 , B. intermedia Noble, 1921 , B. peruana Werner, 1901 , B. talpa Rutven, 1925 , and B. trisanale (Cope, 1868)) , from the B. heteropa group ( B. heteropa ( Lichtenstein, 1956) and B. pallidiceps (Cope, 1862)) (both having hexagonal, smooth dorsal scales), and from B. heteropa and B. flavescens ( B. flavescens (Bonnaterre, 1789) and B. monodactyla (Daudin, 1802)) groups (hindlimbs with more than one digit).
Among the species of B. bresslaui group (diagnostic features for species of B. bresslaui group are summarized in Table 2 View TABLE 2 ), B. geralista sp. nov. can be promptly distinguished from all species by having 30–33 scales around midbody ( 29–30 in B. oxyrhina and 33–47 in the combination of the remaining species); from all species, with exception of B. oxyrhina and B. bresslaui , it differs in having both fore and hindlimbs ending in a apical single scale (forelimbs ending in four apical scales in B. cacerensis Castrillon & Strüssmann, 1998 , hindlimbs with two apical scales in B. micromela , hindlimbs with four clawed toes in B. psamophila , both limbs with four reduced toes and fingers in B. panoplia Thomas, 1965 , and B. scolecoides Vanzolini, 1961 , both limbs with free fingers and toes in B. pyburni Kizirian & McDiarmid, 1998 , and forelimbs with two apical scales in B. didactyla Freitas, Strüssmann, Carvalho, Kawashita-Ribeiro & Mott, 2011 ); from all other species except B. oxyrhina and B. psamophila by the acute, shovel-shaped snout in profile, (snout rhomboid, rounded in profile in B. scolecoides , B. didactyla , B. pyburni , B. cacerensis , B. micromela and rhomboid, slightly protruding upwards in B. bresslaui and B. panoplia ); from all species except B. cacerensis , B. oxyrhina and B. psammophila by the absence of the first temporal, allowing a narrower contact between supralabial and parietal (first temporal present, preventing contact between supralabial and parietal); from all species except B. micromela by a contact between parietal and supralabial (parietal and supralabial without contact), although the contact is with the 6th supralabial in B. geralista sp. nov. and with the 5th supralabial in B. micromela ; from B. panoplia , B. pyburni and B. scolecoides by the absence of prefrontals (present and in contact in P. panoplia and B. pyburni , and widely separated and reduced in size in B. scolecoides ); from B. oxyrhina and B. pyburni by having six supralabials (five and seven, respectively); from B. oxyrhina and B. didactlyla by having the first supralabial and nasal separated (merged); from B. pyburni , B. panoplia and B. micromela by having all ventral scales smooth, quadrangular (lanceolated, keeled in B. pyburni and B. panoplia , and anteriorly quadrangular, smooth, and posteriorly, close to vent, lanceolated, keeled in B. micromela ).
Distribution: Bachia geralista sp. nov. is currently known to occur only in the Planalto dos Gerais, at the eastern Brazilian Cerrado, a large eroded sandstone plateau that covers all western Bahia state, and some parts of Minas Gerais and Tocantins; emerging from the surrounding lowlands, it reaches more than 900 m a.s.l. in some points ( Fig. 4 View FIGURE 4 ). Although we are aware of only a few localities where B. geralista sp. nov occurs, literature records indicates the presence of Bachia at other points of Planalto dos Gerais: Correntina (Colli 1998) and Cocos (Rodrigues et al. 2008), from which specimens not examined. Further examination are need to attest if they belong to this new species.
Natural history: Habitat— Although we sampled all available environments in the area (e.g. dry forest, riparian forest, carrasco and savannas), pitfall trapping at the Peruaçu region, found only three specimens of B. geralista sp. nov., which were found only at the savanna. Active searches within this habitat showed that B. geralista sp. nov. inhabits primarily the leaf litter under larger trees (9 out of 11, found using standardized active search, of which 6 were found under ‘Pequi’ trees ( Caryocar brasiliense ) ( Fig. 5 View FIGURE 5 ). Only one specimen was found under a dead trunk and two under the leaf litter of bushes. Mean abundance was 5 individuals/ha, among all microhabitats, and 6.9 individuals/ha, considering only where lizards were found. Typically, we found single specimens under each patch. Only in two occasions we found more. In one of these situations, we found two large adults and a smaller specimen under an unidentified tree, in the other, four hatchlings within a square meter under a Pequi tree.
Thermal ecology— Temperatures above 40ºC (lethal for B. geralista sp. nov.) were attained at all microhabitats ( Fig. 6 View FIGURE 6 A), in both seasons. Lethal temperatures occurred from 7:00h–16:00h, both in the surface and the exposed subsoil and, only between 13:00–15:00h, under the loose soil covered by vegetation. Temperatures below the fifth percentile (11.38º C) of the frequency distribution of preferred temperatures occurred at all the microhabitats measured. Absolute minimum (7.43ºC) was reached in July (dry season), at 6:00h in the surface of the leaf litter, at the same time, the temperature registered under the loose soil layer was between 14–16ºC. Preferred temperatures of B. geralista sp. nov. were 31.22ºC (50% central range 33.84–28.18ºC) ( Fig. 6 View FIGURE 6 A).
B. oxyrhina B. panoplia B. psamophila B. pyburni B. scolecoides
Prefrontals absent present in contact absent present in contact present separated *Some individuals present seven supralabials.
**Some individuals lack the first temporal, then the 6th supralabial is in contact with parietal. ***Some individuals present two apical scales.
Activity — Although not standardized observations were made, captive specimens, were only seen to be active at hours ranging between 15:00–22:30h.
Behavior— All observed animals, but two, tried to escape using the same tactic in the wild and in both soil types during trials. They first slide quickly over the surface and then burrowed in the loose soil. Another, when found in the wild, jumped a distance of nearly half a meter and entered the leaf litter. A last specimen tried to directly burrow during trials, in both soil types. When grabbed by hand, all the individuals exhibited tail’s prehensile ability.
In the laboratory, one specimen was recorded whipping its tail in an apparently defensive display (http:// www.youtube.com/watch?v=yUi7luTTxlI). Another specimen used its tail as a long support to climb one of its terrarium’s walls ( Fig. 6 View FIGURE 6 B). The four hatchlings found together, were further observed together in three occasions in the laboratory, along the following year, suggesting possible grouping behavior of this species among juveniles ( Fig. 6 View FIGURE 6 C).
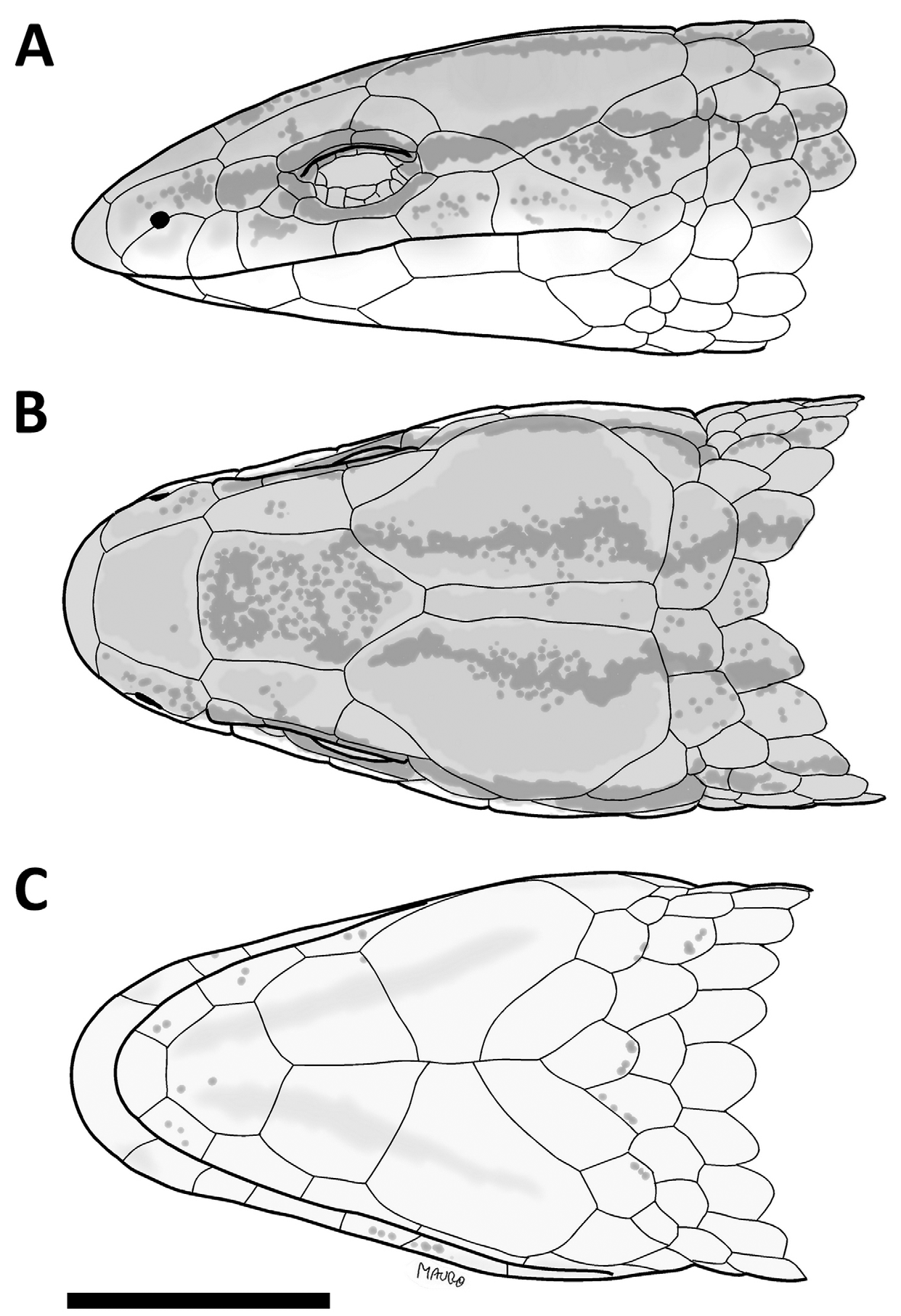

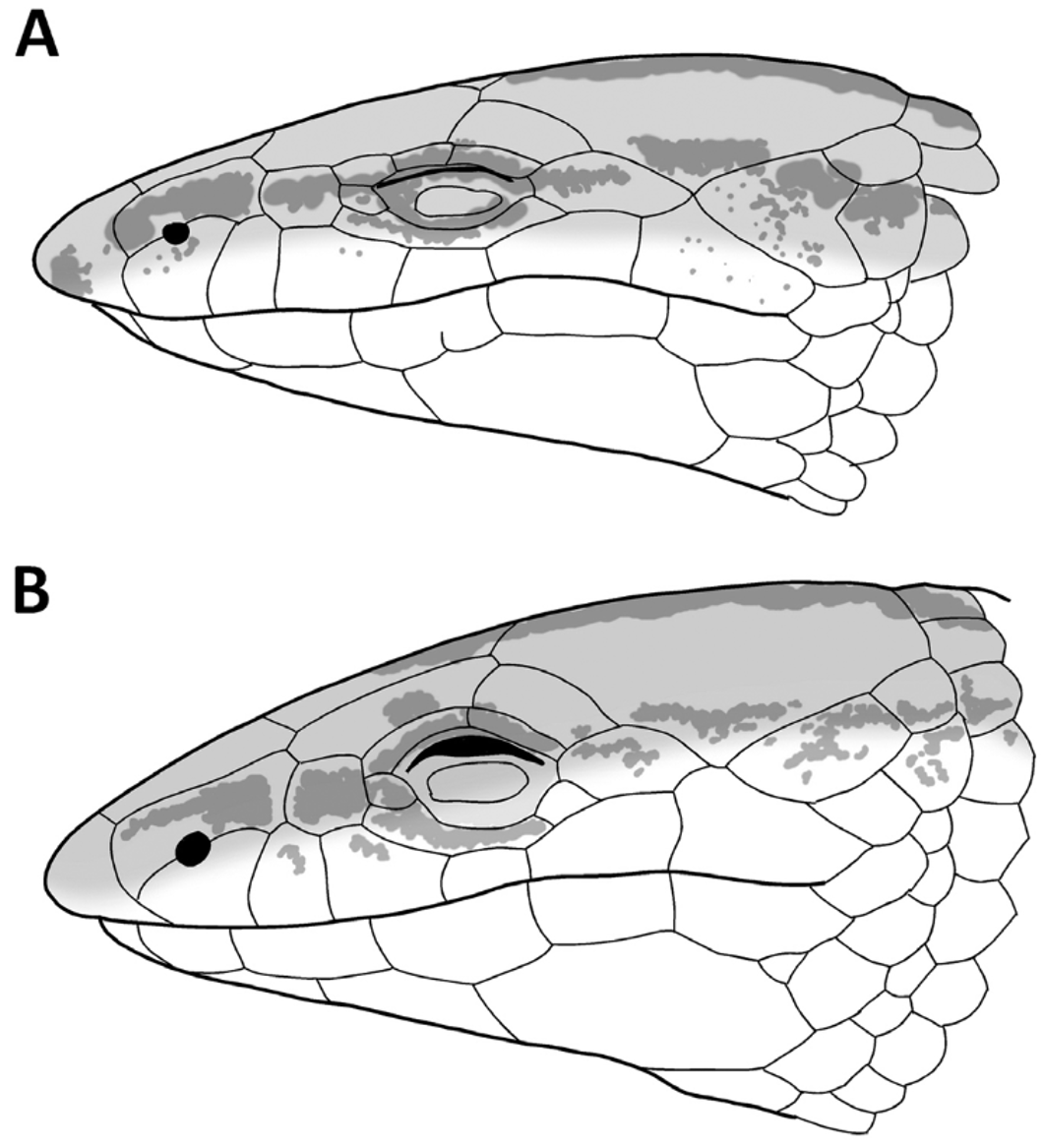
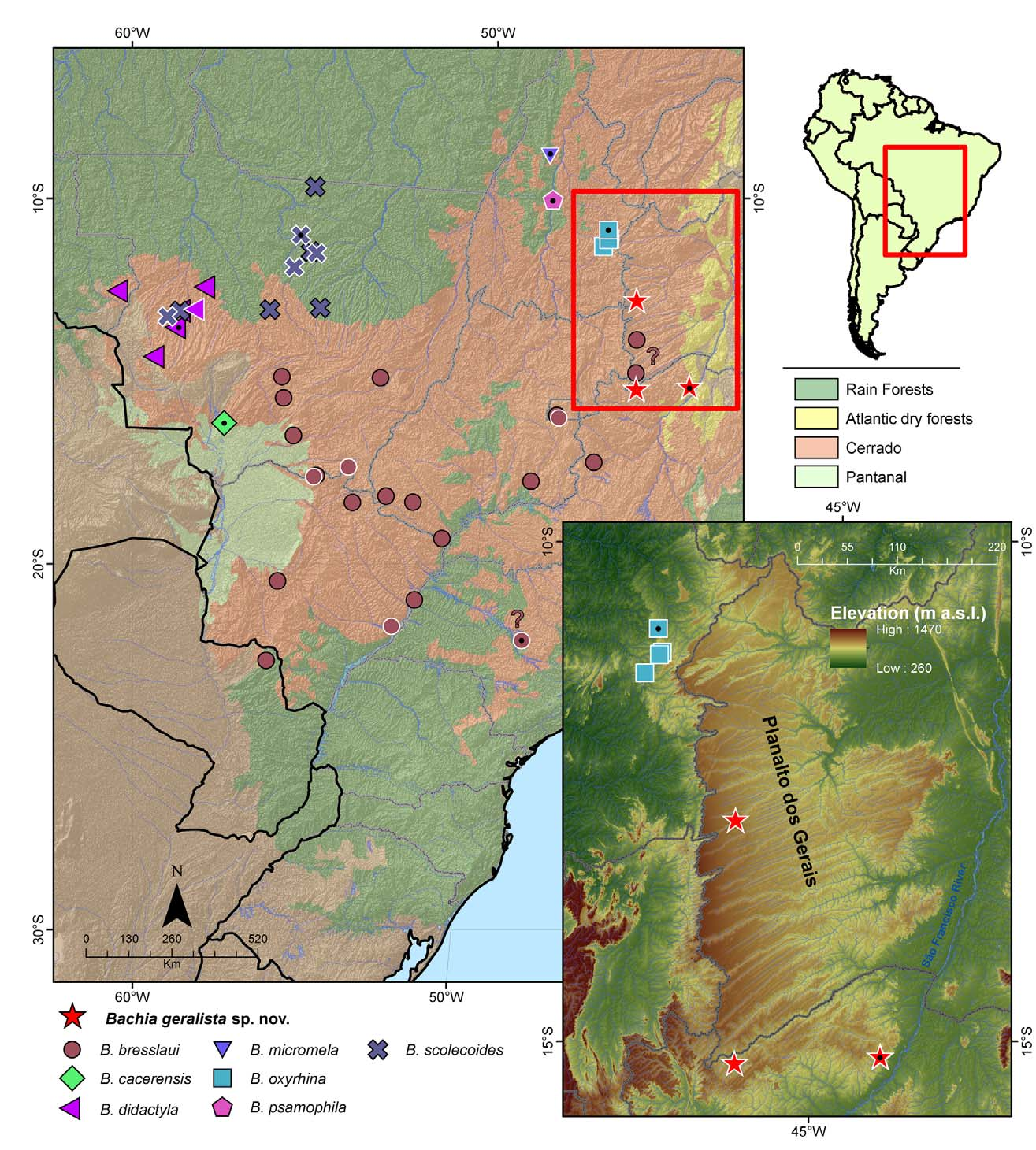

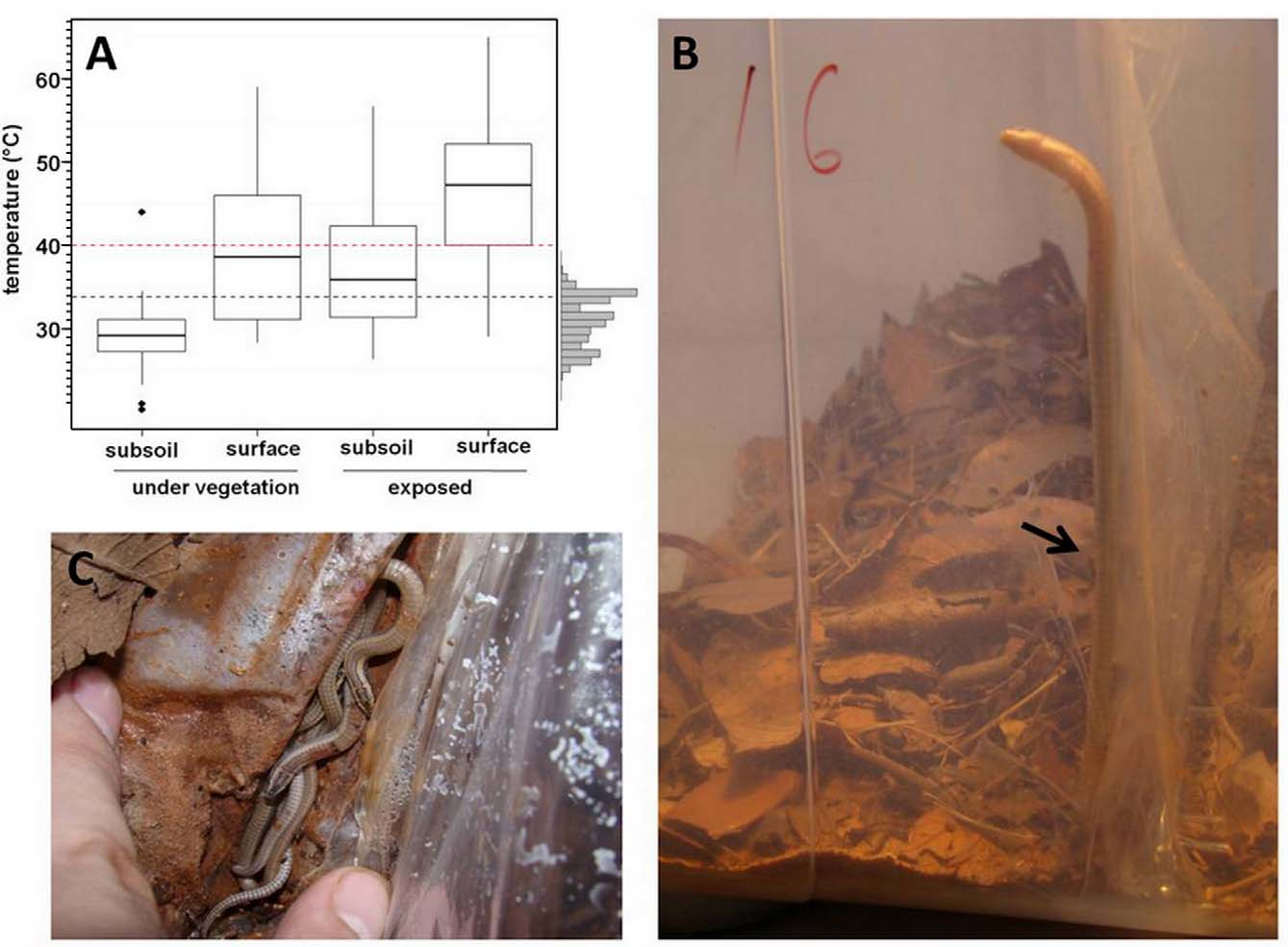
TABLE 1. Meristic and morphometric (mm) values of Bachia geralista sp. nov. and B. bresslaui. Abbreviations are explained in the text.
| SAB | 30–33 | 33–35 SVL | 71.9–101.3 | 64.8–104.2 |
|---|---|---|---|---|
| DO | 46–51 | 47–53 IBNL | 14.6–18 | 14.2–19.2 |
| VE | 36–40 | 36–41 TL | 141–162 | 122.5–148.5* |
| GU | 8–9 | 9–10 LBM | 54.6–79.5 | 48.1–80.8 |
| SL | 6 | 6–7 HH | 3.2–4.6 | 3.5–4.5 |
| CA | 108–115 | 117–123 HW | 4.5–5.8 | 4.6–5.8 |
| PS | 7 | 5–7 HL | 7.6–10.1 | 7.6–10.5 |
| SO | 2 | 2 FLL | 1.8–2.7 | 2–2.8 |
| SC | 2 | 2 HLL | 2–3.9 | 1.9–3.5 |
| FP | 2 | 1–2 END | 3.2–4.2 | 3–4.2 |
| PP | 1 | 1 NND | 2.1–2.8 | 2.1–2.9 |
TABLE 2. Summary of diagnostic features among Bachia species of the B. bresslaui group.
| First supralabial separated and nasal | separated | separated | merged | separated |
|---|---|---|---|---|
| Scales around 30–33 midbody | 33–35 | 35 | 34–37 | 35–38 |
| Contact 6th supralabial supralabial/parietal | no contact** | 6th supralabial | no contact | 6th supralabial |
| First temporal absent | present** | absent | present | absent |
| Contact loreal/ no internasal | no | no | yes | no |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.