
Australolacerta
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3669.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:4A8C9C02-DE18-47B8-8CB7-7F90B89B16AD |
|
DOI |
https://doi.org/10.5281/zenodo.6160987 |
|
persistent identifier |
https://treatment.plazi.org/id/03806730-FF9C-2A49-67EB-FB9BAE67D82C |
|
treatment provided by |
Plazi |
|
scientific name |
Australolacerta |
| status |
|
Monophyly of Australolacerta
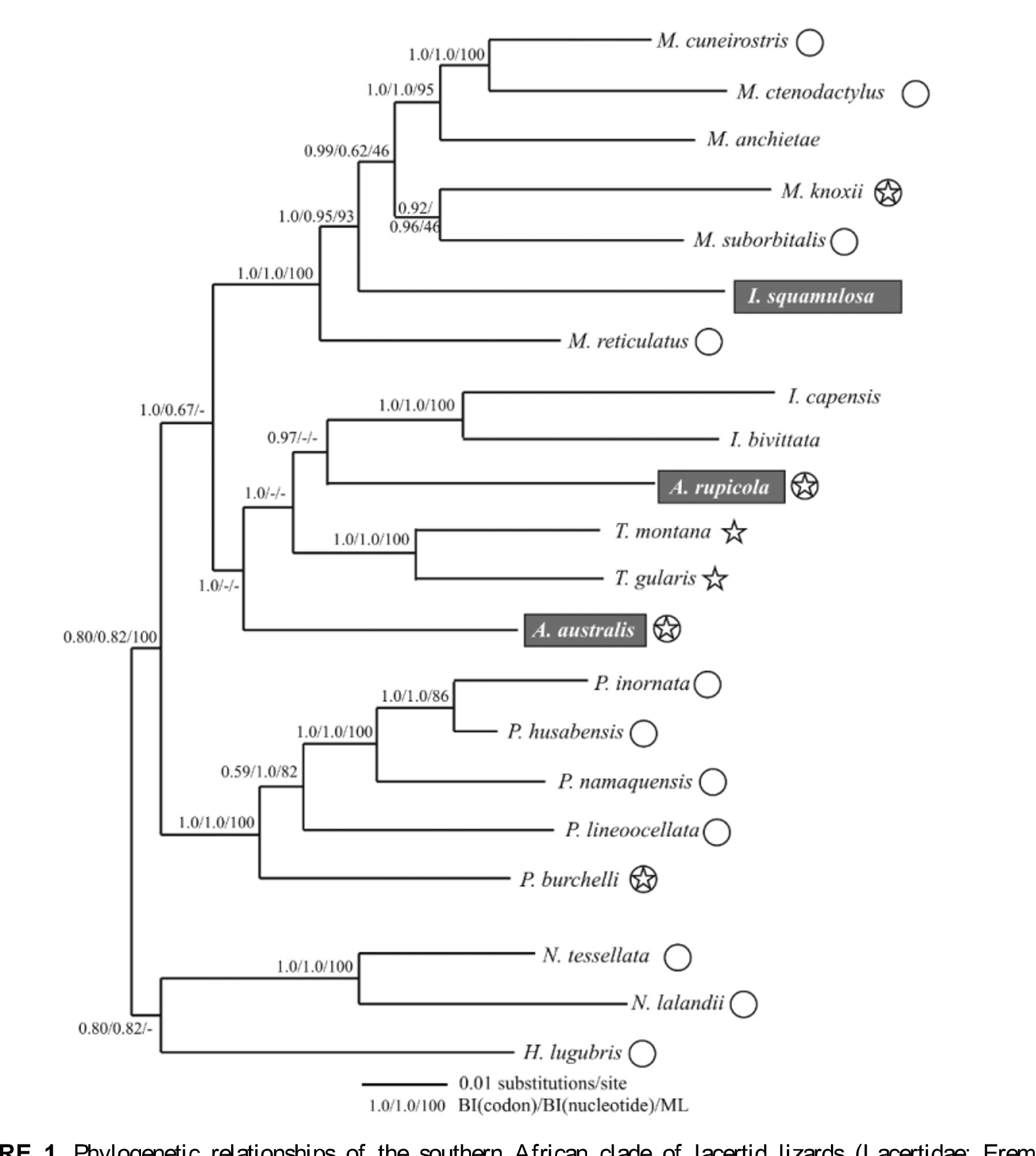
The two species of Australolacerta are endemic to South Africa (Branch 1998), and both were originally placed within Lacerta , creating a zoogeographic paradox as most congeners were restricted to Eurasia (Arnold 1989). Arnold (1989), when describing Australolacerta , gave only a minimal diagnosis, noting that “… the South African species share a number of features with other Ethiopian lacertids which are not found in the apparent closest Palaearctic relatives, namely Lacerta jayakari etc.”. The latter, now transferred to Omanosaura , was initially considered to form a basal lineage within the Eremiadini (Harris et al. 1998), although fuller taxon sampling of African lacertids (Arnold et al. 2007; Hipsley et al. 2009; Kapli et al. 2011) shows it to cluster with a suite of mainly north African genera (e.g. Acanthodactylus , Mesalina , Ophisops ), with Atlantolacerta basal within the Eremiadini (Arnold et al. 2007). The sister relationship of Australolacerta and Tropidosaura proposed by Salvi et al. (2011) and Kapli et al. (2011) was based on the inclusion only of A. australis , and the inclusion of A. rupicola (Edwards et al. 2012; Engleder et al. 2013; this study) revealed the paraphyly of Australolacerta and the basal position of A. australis in a subclade including Ichnotropis , Tropidosaura and A. rupicola .
Both species are rupicolous and Kirchhof and Richter (2009) and Kirchhof et al. (2010a,b; 2012) give details of the species’ biology. They are morphologically similar, albeit that many of these similarities are plesiomorphic within lacertids (Arnold 1989). Due to their high-altitude and small ranges (Branch 1998), the two species have been difficult to collect and therefore little morphological data exists for either species. Recent morphological analyses (Edwards et al. 2012) confirm the similarity between the two species. Yet, important features of hemipenial ornamentation and everted hemipenis structure remain unknown. Whether these similar morphologies reflect adaptive convergence to rupicoly or the retention of plesiomorphic features remains unknown.
The two species are allopatric and geographically separated from one another by a distance of approximately 1700km. Few other genera in southern Africa are known to show such large geographical disjunctions between congeners, and analysis of previous examples has often revealed deep genetic divergence best reflected in generic re-assignment. Examples include: the erection of the genera Kinyongia and Nadzikambia for non-South African dwarf chameleons previously included in Bradypodion (Tilbury et al. 2006) , and Inyokia for the problematic Swazi rock snake that was shown to be sister to the tropical forest snake Homonotus modestus (Kelly et al. 2011) . In one of the few exceptions of congeneric range disjunctions within the subcontinent, cordylid flat lizards of the Platysaurus capensis complex are separated geographically from other Platysaurus by approximately 850km (Branch & Whiting 1997). Other described lacertid species are also separated from congeners by large distances, for example Heliobolus lugubris is separated from its congeners ( H. spekii , and H. nitidus ) by> 2000km and Ichnotropis chapini is separated from other Ichnotropis by approximately 2000km (Branch, 1998; Spawls et al. 2002). However, the Central African region is undersampled and it is possible that with increased sample collection new species may be discovered or that ranges of described species may increase, lessening the geographic gap between congeners.
Although there were no significant differences between the obtained trees and the constrained trees in the SH or AU test when the topology was constrained to monophyly for Australolacerta sequence divergence estimates and the long branch lengths in the phylogenetic analyses ( Figure 1 View FIGURE 1 ) all other evidence strongly suggests that the two Australolacerta species do not share a recent evolutionary history (Edwards et al. 2012).To provide consistency between taxonomic divisions in the Eremiadini, we propose that the two species of Australolacerta should be placed in separate genera. The type species of Australolacerta is Lacerta australis (Arnold 1989) , and we therefore erect a new genus for the remaining species Lacerta rupicola , based on morphology and genetic divergence.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |