
Atlantisina tricornis, Berning & Harmelin & Bader & Cibio, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.347 |
|
DOI |
https://doi.org/10.5281/zenodo.3850622 |
|
persistent identifier |
https://treatment.plazi.org/id/546F87A1-FFAE-FF83-094B-93273555F825 |
|
treatment provided by |
Carolina |
|
scientific name |
Atlantisina tricornis |
| status |
gen. et sp. nov. |
Atlantisina tricornis gen. et sp. nov.
urn:lsid:zoobank.org:act:0DB29D39-154D-4C9D-AF4D-F07E525B850B
Figs 1E View Fig , 6 View Fig A–F, Tables 6–7
Hippoporina View in CoL sp. – d’Hondt 1974: 46, fig. 6.
Romancheinidae View in CoL gen. et sp. indet. – Souto et al. 2016: 432.
Diagnosis
Frontal shield with a reticulate pattern of raised ridges around polygonal depressions; aperture with six oral spines; orifice suborbicular, proximal edge fairly straight or slightly concave, condyles short, blunt, tip somewhat thickened; suboral area with three tall thick mucrones with pointed tips, occasionally branching from their base or with bifid tips, most often with a central, vertically oriented mucro that is occasionally paired, and two mucrones proximolateral to orifice that are slightly curved and point outwards or are vertically oriented. Ooecium as long as wide or occasionally laterally compressed; ectooecium well developed, approximately covering two-thirds or more of entire ooecium; exposed endooecium relatively small, occasionally reduced to an elongated central area, surface topography similar to that of frontal shield, but usually with smaller depressions and steeper ridges. Ancestrula with nine spines, the distal four more closely spaced than (and slightly offset from) the proximal five spines, opesia slightly constricted in distal fourth, cryptocyst practically absent.
Etymology
Named for its prominent tridentate suboral mucro.
Material examined
Holotype
N IBERIAN SLOPE: 1 large ovicellate colony on biogenic substratum (plus another smaller colony), Stn 39 ( MNHN-IB-2014-60 ).
Paratypes
N IBERIAN SLOPE: 2 colonies on Lophelia pertusa (Linnaeus, 1758) , Stn 38 (MNHN-IB- 2014-61); 5 colonies on a fragment of coral skeleton, Stn 39 (MNHN-IB- 2014-63); 1 coated ovicellate colony with ancestrula, Stn 39 (MNHN-IB- 2014-64); 1 coated ovicellate colony, Stn 39 (MNHN-IB- 2014-65).
LE DANOIS BANK:1 colony on Balanophyllia thalassae Zibrowius, 1980 ,Stn 35 (MNHN-IB- 2014-62).
GALICIA BANK: 1 coated colony, Stn 15 (MNHN-IB-2014-279).
Other material examined
N IBERIAN SLOPE: 2 colonies on old L. pertusa skeleton, Stn 37 (unregistered MNHN material); 2 colonies on coral skeletons, Stn 39 (unregistered MNHN material); 2 colonies on coral skeleton, Stn 40 (unregistered MNHN material); 15 colonies on fragmented coral skeletons, Stn 39 ( OLL 2016/145).
GALICIA BANK: several colonies identified as Romancheinidae gen. et sp. indet. by Souto et al. (2016), Stn 13 ( MNCN 25.03/3955); 1 colony on coral, Stn 14 (unregistered MNHN material); 4 colonies on rock, Stn 15 (unregistered MNHN material); 3 colonies on rock, Stn 16 (unregistered MNHN material).
W IBERIAN SLOPE: 2 colonies, identified as Hippoporina sp. by d’Hondt (1974), Stn 44 (MNHN- IB-2008-7194).
Description
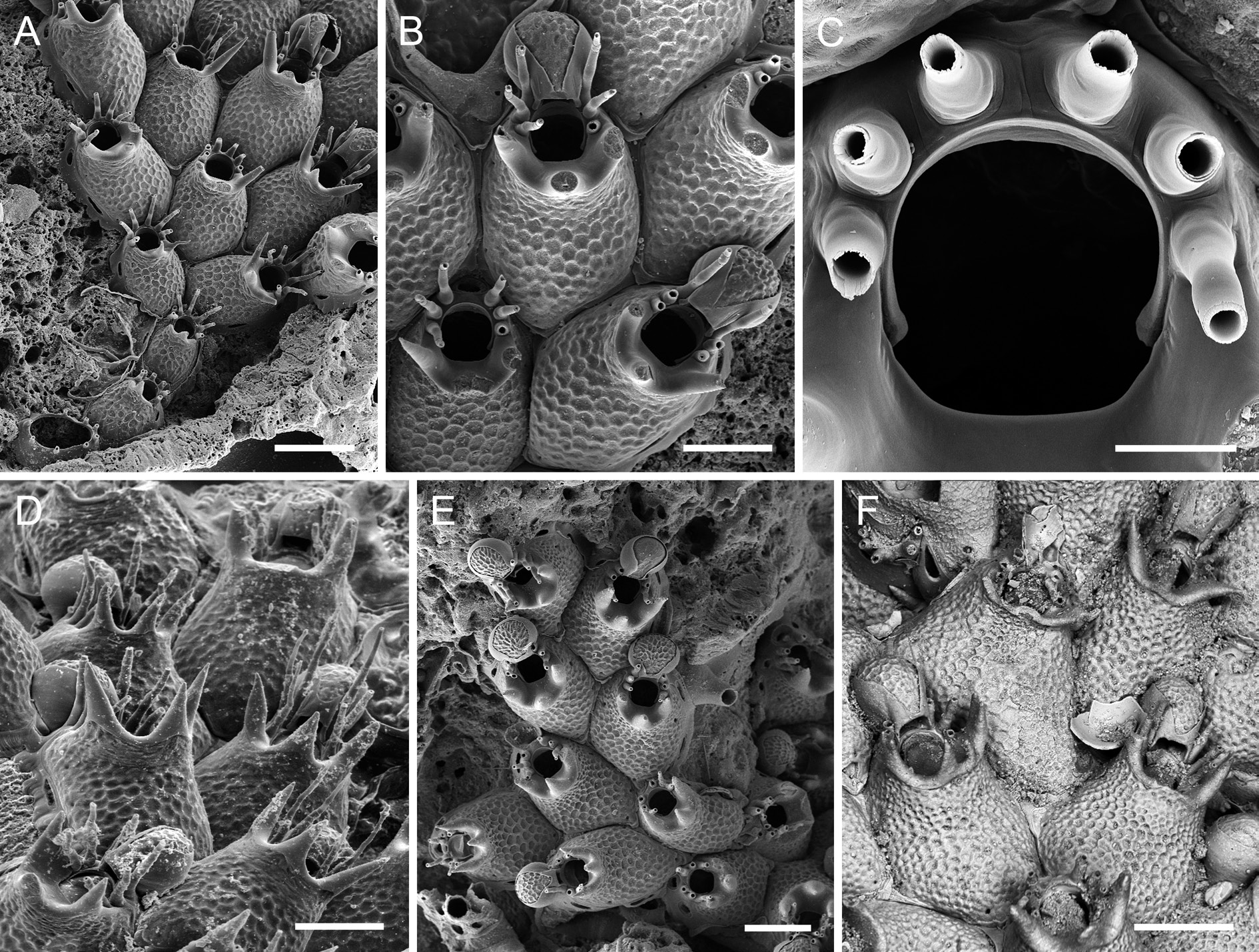
Colony encrusting, unilaminar, forming small patches and bi- to triserial ribbons ( Fig. 6E View Fig ). Zooecia polygonal or oval, with tapering proximal end(s) wedged in between proximal zooecia, separated by deep grooves ( Fig. 6 View Fig A–B). Frontal shield convex, distally steeply raised to form a suboral crest, surface with a reticulate pattern of raised ridges around polygonal depressions ( Fig. 6B View Fig ), imperforate except for a few small marginal pores hardly visible in frontal view or in older zooecia; suboral crest usually with three widely spaced and thickly calcified conical mucrones with acuminate tips ( Fig. 6D View Fig ), the mucrones either directed vertically ( Fig. 6E View Fig ) or diverging outwards ( Fig. 6F View Fig ), occasionally one or all mucrones have bifid tips and/or there may be a pair of central mucrones ( Fig. 6F View Fig ); all mucrones rising from a prominent broad area of smooth gymnocystal calcification that slopes towards proximal orifice margin, then narrowing distally and abutting proximal pair of spines; lateral wall moderately well developed, septular pores transversely oval to elongate, area surrounding pores reduced, distal pore large, suborbicular.
Orifice suborbicular, proximal border fairly straight to slightly concave, widest in distal third, proximal third delimited by a pair of very short blunt condyles with slightly thickened tips ( Fig. 6C View Fig ); lateral and distolateral margins with six spines arranged in two series of three, separated by a distinct distal gap ( Fig. 6D View Fig ), spines up to some 350 µm long, comprising a thick tubular base (80–85 µm high) and a thinner whip-like part (200–260 µm long); all 6 spines present in ovicellate zooids, with distal pair abutting proximolateral ooecial wall and wedging in ooecial aperture on both sides.
Ovicell hyperstomial, ooecium barely resting on frontal shield of distal zooid with its proximal part or entirely free at colony margin, globular, about as long as wide or laterally compressed, with a short tubular peristome terminating at distal apertural margin ( Fig. 6B View Fig , E–F); ectooecium well developed, covering about two-thirds or more of entire ooecium; exposed endooecium of variable size and shape, either pear-shaped and narrowing on peristome when large, or forming a broad central strip when ooecium is compressed, surface topography similar to that of frontal shield but with smaller and marginally elongate depressions and steeper ridges; ooecial aperture about as tall as wide, acleithral.
Ancestrula oval (ca 350 µm long, 230 µm wide), smooth gymnocyst narrow all around, cryptocyst virtually absent; opesia large, oval to pyriform (ca 235 µm long, 145 µm wide), slightly constricted in distal fourth; nine spines, with five proximal ones widely spaced and four distal ones situated closer together ( Figs 1E View Fig , 6A View Fig ).
Remarks
Atlantisina tricornis gen. et sp. nov. is the only species of Atlantisina gen. nov. that was recorded from the continental slope while all other species occur on seamounts and near islands. Moreover, it is the only species of Atlantisina gen. nov. that has previously been recorded and figured (d’Hondt 1974; Souto et al. 2016).
Atlantisina tricornis gen. et sp. nov. is clearly distinguished from all other congenerics by its thickly calcified, suboral crest with three pointed mucrones. However, there is some variability in mucro shape and orientation between colonies from the same area, as well as between populations from the three sampled regions, the northern Iberian slope, Galicia Bank and the western Iberian slope. For instance, in the Galicia Bank population the three mucrones are usually single, unbranched, straight, and vertically oriented ( Fig. 6E View Fig ), whereas in some zooids the lateral mucrones may be slightly curved outwards and branched, and the central mucro may be twinned. Branching of mucrones was rather frequent in colonies from the continental shelf off northern Portugal (W Iberian slope; Fig. 6F View Fig ), while in the colonies from the N Iberian slope the lateral mucrones were constantly unbranched and diverging slightly outwards ( Fig. 6 View Fig A–B, D). However, in some colonies from this region, a secondary acuminate tip may occur laterally on the central mucro. Thus, although the end-members of the mucro-morphotypes are distinctly different, a clear distinction between regions cannot be drawn, as intermediate stages are present.
A certain variability was also detected in the development of the ooecium, whose shape ranges from globular with a large suborbicular area of exposed endooecium ( Fig. 6E View Fig ) to laterally compressed ones in which the endooecium is reduced to a narrow central area ( Figs 6B, F View Fig ). Consequently, ooecium width and the length-width ratio (OvL/OvW) may differ between colonies. In order to assess the morphological variability between colonies and regions, one-way ANOVA was were performed on the original length- and width-measurements of zooids, orifices and ooecia. As morphological differences were optically noticeable between colonies occurring in Galicia Bank, the populations were divided into four areagroups, the N Iberian slope (NIS), Galicia Bank 1 (GAL1), Galicia Bank 2 (GAL2), and W Iberian slope (WIS). Statistical analyses of ooecium width data show that, while the mean values are similar within two area-pairs, ooecium width is significantly higher in WIS and GAL1 than in NIS and GAL2 (F = 24.35, p <0.001; see Table 7 View Table 7 ). Mean values of ooecium length/width ratio of GAL1 (0.98) are significantly different from NIS (1.32), while similar intermediate values (1.15) are observed in WIS and GAL2 colonies.Although distinct differences in size of autozooids and orifices can be perceived between colonies from the four sampled areas ( Table 7 View Table 7 ), statistical analyses do not show a common hierarchy in the four populations when considering the different dimensions (ZL, ZW, OL, OW). Thus, concerning both morphology and morphometry, there is no clear distinction between the populations of the three geographic areas that would allow separating them at the species level.
With their bifid tips and twinned central mucrones ( Fig. 6F View Fig ), some of the zooids in the Portuguese morphotype of A. tricornis gen. et sp. nov. somewhat resemble Atlantisina lionensis gen. et sp. nov. from Lion and Seine seamounts (see below). However, in the latter the mucrones are not as thickly calcified, are positioned on a distinctly raised and relatively straight crest, and the lateral mucrones point distally.
Calcification of the frontal shield’s surface in A. tricornis gen. et sp. nov. and the other new species introduced below is the exact opposite of that of A. atlantis gen. et sp. nov., A. meteor gen. et sp. nov. and A. seinensis gen. et sp. nov. described above. With the reticulate pattern of ridges delimiting round to hexagonal depressions in the former group ( Fig. 6B View Fig ), the precipitation of carbonate seems to be the negative blueprint of the frontally flattened, round to polygonal nodules bounded by grooves in the latter group.
Ecology
Atlantisina tricornis gen. et sp. nov. has been found at depths between 450 and 1040 m on the continental slope, and between 675 and 1700 m on Galicia Bank (see also Souto et al. 2016: table 22, listed as “Species indet.”). The species forms small patches and bi- to triserial colonies that encrust rocks and biogenic substrata, mainly coral skeletons but also brachiopods, balanid plates and cidarid spines.
Distribution
Atlantisina tricornis gen. et sp. nov. was recovered from the continental slope of northern to western Iberia as well as from Galicia Bank.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Neocheilostomina |
|
SuperFamily |
Lepralielloidea |
|
Family |
|
|
Genus |
Atlantisina tricornis
| Berning, Björn, Harmelin, Jean-Georges & Bader, Beate 2017 |
Romancheinidae
| Souto J. & Berning B. & Ostrovsky A. N. 2016: 432 |
Hippoporina
| d’Hondt 1974: 46 |