
Anthystrix Kirejtshuk, 1981
|
publication ID |
https://doi.org/ 10.5281/zenodo.5319334 |
|
DOI |
https://doi.org/10.5281/zenodo.10542373 |
|
persistent identifier |
https://treatment.plazi.org/id/03BE87CC-F634-FFE5-BAC4-FB9FFC20F9A8 |
|
treatment provided by |
Felipe |
|
scientific name |
Anthystrix Kirejtshuk, 1981 |
| status |
|
26. Anthystrix Kirejtshuk, 1981
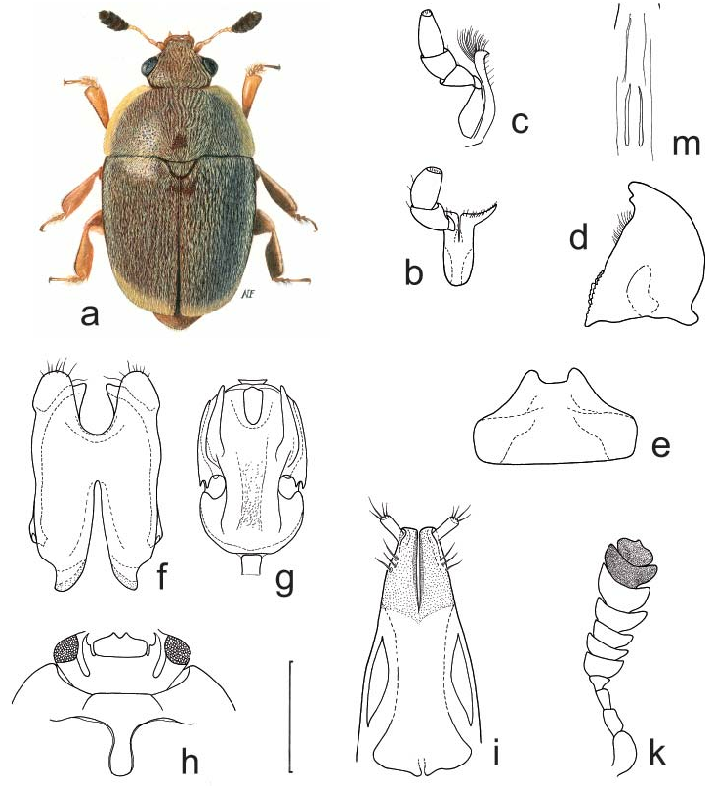
( Figs. 26 a–m View Fig )
Anthystrix Kirejtshuk, 1981: 133 .
Type species. Pria squamosa Grouvelle, 1899: 146 (by original designation) [= Anthystrix squamosa (Grouvelle, 1899) ].
Generic redescription and diagnosis. Inclusive species vary moderately in size (2.0– 2.7 mm length), and share the following combination of characters.
Body color and pubescence: pubescence always long, golden to silvery-whitish and dense, recumbent, partially obscuring the predominantly dark brown dorsal body surface, not always obscuring the narrowly flattened and frequently pale pronotal and elytral sides ( Fig. 26a View Fig ); lateral margin of pronutum and elytra with a series of faintly distinct, small and short setae, each seta 0.3–0.5× as long as those on elytral disc; posterior margin of pronotum
3) Recently examined material in NHMB collections (AUDISIO unpublished data).
with relatively short, distally multifid microsetae, also present on middle portion anterior to scutellum (as in Fig. 27f View Fig ).
Dorsal habitus: body moderately convex, oval ( Fig. 26a View Fig ; Figs. 1–5 View Fig View Fig View Fig View Fig View Fig in AUDISIO et al. 2009a); dorsal punctures on discal portion of pronotum as large as or larger than eye facet, moderately to deeply impressed ( Fig. 26a View Fig ); anterior margin of clypeus medially truncate, simple, i.e. without small distinct medial bulge, and usually distinctly bordered ( Fig. 26a View Fig ); circum-ocular furrows (occipital sulci) on head deeply impressed and complete (as in Fig. 27i View Fig ); eyes large and usually markedly projecting laterally ( Fig. 26a View Fig ); posterior angles of pronotum distinct, blunt, obtuse, never directed posteriorly ( Fig. 26a View Fig ); scutellum uniformly punctured on most of exposed portion (as in Fig. 27f View Fig ); elytra with simple punctures, never transversely strigose; elytral humeral angle faintly distinct, widely obtuse, never protruding laterally ( Fig. 26a View Fig ); elytral humeral striae indistinct; elytral pre-sutural striae faintly visible, originating at scutellar vertex or slightly posterior, terminating at elytral apex, and delimiting on each elytron a faintly distinct, flat, unraised sutural border, more distinct at distal fourth, narrower than proximal width of 3 rd antennomere; elytral apices truncately rounded in both sexes ( Fig. 26a View Fig ); pygidium partially exposed, moderately convex, apically rounded in both sexes ( Fig. 26a View Fig ).
Ventral habitus: antennal furrows markedly delimited, moderately convergent posteriorly; mentum subpentagonal ( Figs. 26e, h View Fig ); prosternal antennal furrows on anterior margin of prosternum strongly delimited and raised, distinctly divergent posteriorly, never extending posteriorly to anterior margin of procoxal cavity ( Fig. 26h View Fig ); prosternal process moderately wide, subapical portion 1.9–2.1× as wide as maximum width of 1 st antennomere, apex usually bluntly rounded ( Fig. 26h View Fig ); lateral borders of prosternal process delimiting shallowly impressed and faintly distinct furrows, never distally terminating over predistal lateral expansions (as in Fig. 27g View Fig ); posterior margin of mesoventrite simple, never medially incised; moderate to marked sexual dimorphism in impressions on metaventrite; first two visible abdominal ventrites simple in both sexes, without tufts of setae, caudal marginal lines of metacoxal cavities simple, subparallel and contiguous to posterior margin of metacoxal cavities, with moderately deep arched impression of outer ‘axillary’ line; ‘axillary’ space on first abdominal ventrite reduced, ‘axillary’ angle nearly right angled; strongly marked and deeply impressed arched impressions on basal portion of last visible abdominal ventrite, only partially covered by distal portion of penultimate visible abdominal ventrite (as in Fig. 27g View Fig ).
Appendages: male 1 st antennomere 0.8–1.0× as long as width of protibiae excluding distal teeth ( Figs. 26a, k View Fig ); 3 rd antennomere usually 2× as long as wide in both sexes, nearly as long as and slightly thinner than 2 nd; 4 th and 5 th antennomeres subequal in females, moderately short and slightly longer than wide, but strongly modified in males when associated with antennal club ( Fig. 26k View Fig ); male antennal club compact, comprising last 4 to 9 antennomeres, spectacular but apparently never allometric development ( Figs. 26a, k View Fig ), 3-jointed in females; labial palpi relatively short in both sexes ( Fig. 26b View Fig ), terminal segment 1.5–1.7× as long as wide; maxillary palpi relative long and slender in both sexes ( Fig. 26c View Fig ), terminal segment 1.8–2.0× as long as wide; mandible mid-sized, moderately short, apex bifid, moderately acuminate, sexual dimorphism absent ( Fig. 26d View Fig ); tarsi of normal size and shape, 0.7–0.9× as long as corresponding tibiae ( Fig. 26a View Fig ); tarsal claws simple, never toothed at base (as in Fig. 24f View Fig ); protibiae usually with reduced teeth on outer margins ( Fig. 26a View Fig ); lateral margin of meso- and metatibiae bearing a single and regular row of long, thin, yellowish pegs ( Fig. 26a View Fig ), without U-shaped sinuosity at distal third; meso- and metatibiae variably shaped, long and slender to wider and shorter, subtrapezoidal ( Fig. 26a View Fig ; Figs. 1–5 View Fig View Fig View Fig View Fig View Fig in AUDISIO et al. 2009a); sexual dimorphism variably developed in meso- and metatibiae (rarely protibiae); tarsal plates of prolegs more or less distinctly wider in males; posterior margin of metafemora simple in both sexes, without tubercles or projections.
Male genitalia: peculiarly shaped, with more or less developed and characteristic processes along inner side of parameres ( Fig. 26f View Fig ), and with deep median longitudinal desclerotization from proximal portion of tegmen extending to medial distal V-shaped excision; median lobe of aedeagus violin-shaped, with strong lateral emargination, and peculiarly shaped cup-like distal projection ( Fig. 26g View Fig ); main sclerites of internal sac (flagellum) small, linear, and lightly sclerotized, typically 4–5× shorter than aedeagus ( Fig. 26m View Fig ).
Female genitalia (ovipositor): uniformly shaped, large; styli usually long and distinct, simple, unpigmented, inserted near to apex of contiguous gonostyloids, each gonostyloid lightly sclerotized and moderately pigmented distally, with a simple, never indentate outer portion of basicoxites ( Fig. 26i View Fig ), and a single, small, pigmented and more sclerotized arcuate area along outer subdistal portion of gonostyloids. ‘Central point’ of ovipositor usually located more distad than middle, without proximad directed spicule.
Etymology. KIREJTSHUK (1981) did not specify the etymology of his newly described genus Anthystrix . The generic name is obviously derived from Greek ‘υστριξ’ (hystrix in Latin) meaning a hedgehog, which is indicative of the long, spine-like, and colored setae covering most of the body surface, and is likely combined with the Greek suffix ‘αντί’, also meaning ‘similar to’, or ‘to be compared with’. The gender of this genus was implicitly (but erroneously) considered to be masculine in the original description ( KIREJTSHUK 1981), but was later correctly considered feminine ( KIREJTSHUK & EASTON 1988) according to the recognized feminine gender of Hystrix Linnaeus, 1758 ( Mammalia, Rodentia ; the Old World porcupines).
Biology. All species are apparently strictly associated for larval development with male inflorescences of Asteraceae within the phylogenetically isolated tribe Tarchonantheae , in particular with Tarchonanthus L. ( AUDISIO et al. 2008, 2009a).
Phylogenetic position. See the above discussion regarding the phylogenetic position of Chromogethes and members of the ‘ Anthystrix complex of genera’. With regards to Anthystrix sensu stricto, this genus exhibits a series of autapomorphic chararcters, listed above in the generic diagnosis, which clearly suggests a relatively isolated phylogenetic position in the [‘ Anthystrix complex of genera’ + ( Chromogethes )] clade. The peculiar antennal morphology of Anthystrix species is likely a recent product of an evolutionary radiation mainly driven by sexual selection (KODRIC- BROWN et al. 2006), and associated with hyperthelic development of antennal characters in males, which is unusual in members of the Meligethes -complex of genera ( JELÍNEK 2000a).
Taxonomy and geographic distribution. This taxon includes six southern African species, attributed to three formerly recognized species-groups, i.e. the ‘ squamosa ’, ‘ longiclava ’, and ‘ endroedyi ’ species-groups ( AUDISIO et al. 2009a).
Anthystrix endroedyi Audisio & Cline, 2009 South Africa: Kwa Zulu-Natal, NW Region (= Anthystrix martini auct., nec Grouvelle, 1899)
Anthystrix flabellicornis Audisio & Cline, 2009 South Africa: E Western Cape
Anthystrix longiclava Kirejtshuk & Easton, 1988 South Africa: Eastern Cape
Anthystrix megalocera Audisio & Cline, 2009 South Africa: E Western Cape
Anthystrix nigroclava Kirejtshuk & Easton, 1988 South Africa: SE Western Cape
Anthystrix squamosa (Grouvelle, 1899) South Africa: SW Western Cape







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Anthystrix Kirejtshuk, 1981
| Audisio, Paolo, Cline, Andrew Richard, Biase, Alessio De, Antonini, Gloria, Mancini, Emiliano, Trizzino, Marco, Costantini, Lorenzo, Strika, Sirio, Lamanna, Francesco & Cerretti, Pierfilippo 2009 |
Anthystrix
| KIREJTSHUK A. G. 1981: 133 |