
Ampharete cirrata Webster & Benedict, 1887
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5174.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:43181489-8314-40B1-B451-F950B7872EC6 |
|
DOI |
https://doi.org/10.5281/zenodo.6992814 |
|
persistent identifier |
https://treatment.plazi.org/id/2F1987D9-704C-8772-70AA-891CFF33F8F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Ampharete cirrata Webster & Benedict, 1887 |
| status |
|
Ampharete cirrata Webster & Benedict, 1887 View in CoL
( Figs 4–8 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Ampharete cirrata Webster & Benedict, 1887: 748 View in CoL , Pl. VIII, Figs 110–112.
Diagnosis. Medium-sized species of up to 21 mm in length and 2.8 mm in width. Twelve thoracic, two intermediate, and ten abdominal uncinigers with very long neuropodial cirri. Branchiae arranged in two groups separated by a median gap of about 1.5 branchial base width; three pairs in anterior transverse row and last pair in posterior position, directly below middle branchia of anterior transverse row. Paleae long and thin with filiform tips, 9–18 each side. One pair of nephridial papillae on segment IV. Pygidium with up to 17 anal cirri, arranged in two (more or less distinct) rows.
Material examined. Type material (based on photographs): North Atlantic Ocean , Eastport, Maine ( USA). Syntypes of Ampharete cirrata Webster & Benedict, 1887 , USNM 457 About USNM (3 specimens) .
Non-type material: Baltic Sea, north from Isle of Rügen. Site 1–5 ( Table 1 View TABLE 1 ), SBRO-P 95 (23 specimens) , SBRO-P 5528 (5 specimens) , SBRO-P 17367 (134 specimens) , ZSRO-P2669 (16 specimens) , ZSRO-P2670 (15 specimens) .
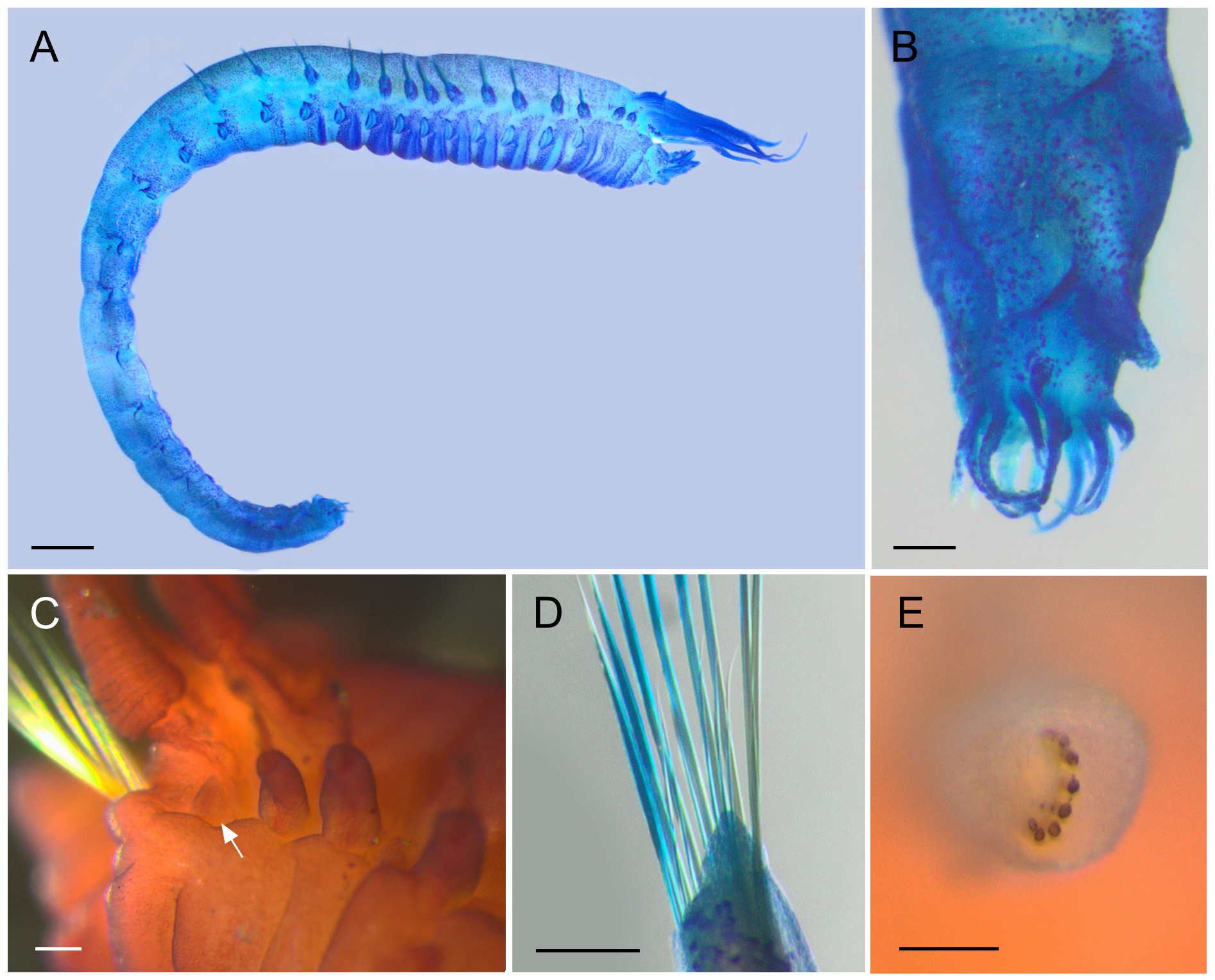
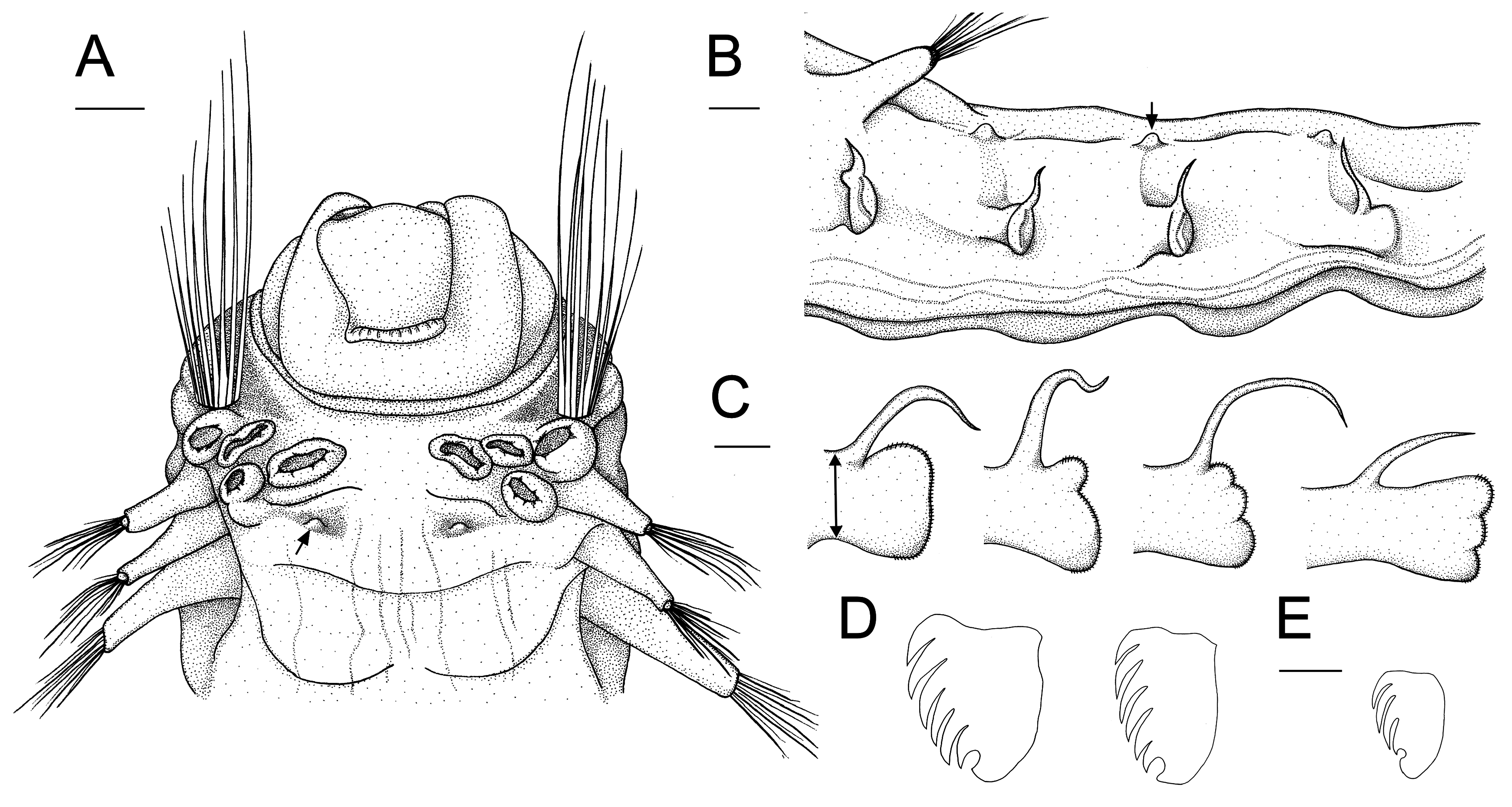
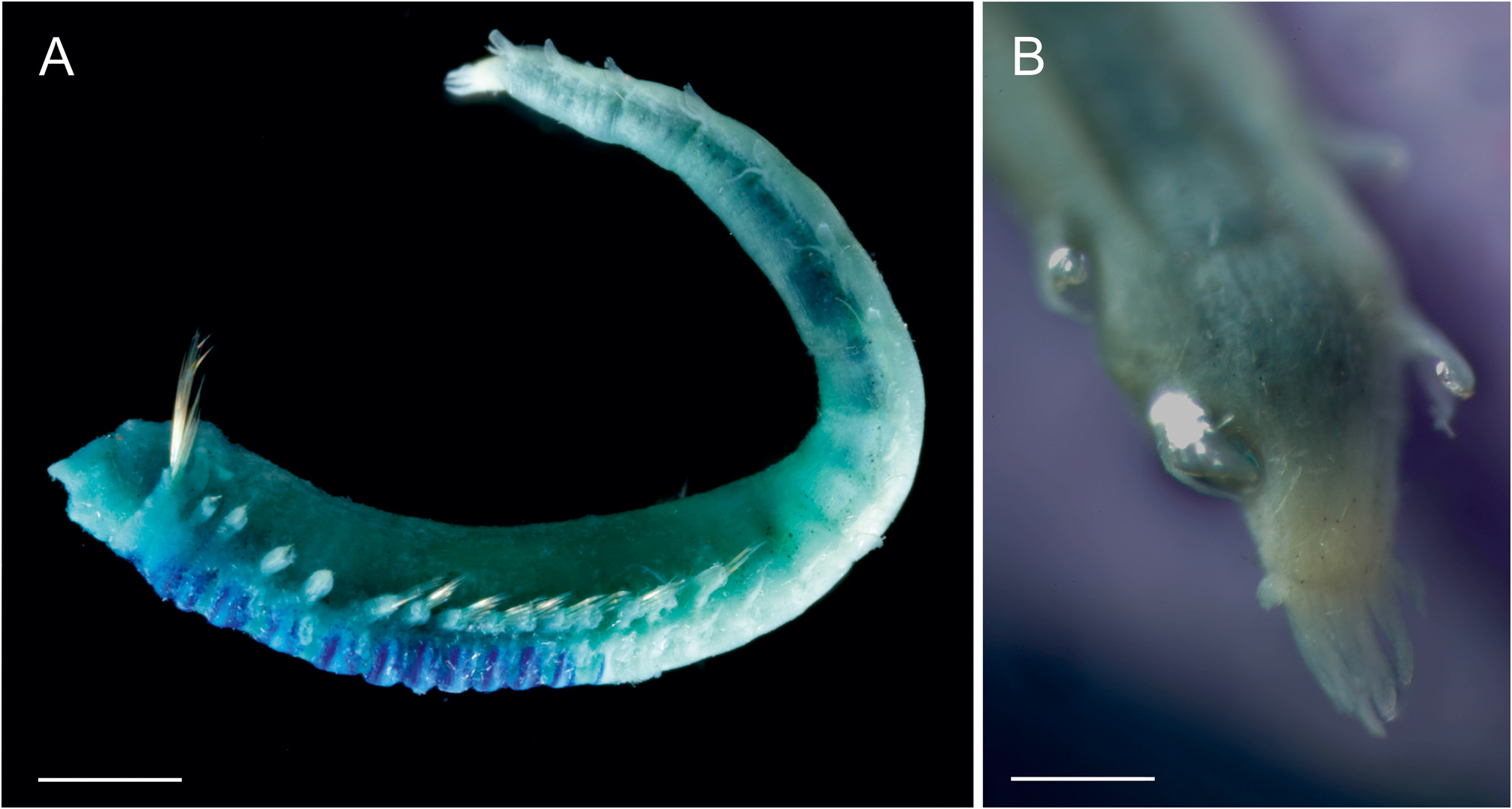
Description. Complete specimens 11–21 mm long and 1.7–2.8 mm wide in thorax ( Fig. 4A View FIGURE 4 ). Prostomium trilobed; median lobe U-shaped and separated by deep lateral grooves ( Figs 5A View FIGURE 5 , 6B,E View FIGURE 6 ); prostomial glandular ridges and eyespots absent. Nuchal organs not observed. Buccal tentacles papillose ( Fig. 6A,C View FIGURE 6 ). Four pairs of branchiae arranged in two distinct groups with a median gap of about one and a half times the width of branchial bases ( Fig. 5A View FIGURE 5 ); branchiae not fused at base; branchiae thick at base and gradually tapering towards distal end, longest branchiae reaching thoracic chaetiger 5–9 ( Figs 4A View FIGURE 4 , 6E View FIGURE 6 ); anterior three pairs of branchiae arranged in a transverse row on segment II/III, fourth pair slightly posterior to anterior row, directly below middle branchia of transverse row ( Fig. 5A View FIGURE 5 , branchial bases can be slightly deformed and displaced through fixation); branchiae of segment II in 2nd outermost position of anterior row, branchiae of segment III in outermost position of anterior row, branchiae of segment IV in innermost position of anterior row, branchiae of segment V in posterior position. One pair of nephridial papillae, dorsally between the two innermost branchiae on segment IV ( Figs 5A View FIGURE 5 , 6E View FIGURE 6 ). Segment II with 9–18 long, thin and slender paleae on each side, gradually tapering into filiform tips ( Figs 5A View FIGURE 5 (only drawn 8 paleae), 6A,B,D,E); paleae clearly protruding beyond the prostomium, at least two times as long as middle lobe of prostomium ( Fig. 5A View FIGURE 5 ); arranged in a semicircle, with a few small thin paleae on the outer edge of the semicircle and longer thick paleae close to prostomium ( Fig. 6D View FIGURE 6 ). Thorax and abdomen of about similar length; thorax slightly wider than abdomen; abdomen tapering posteriorly ( Fig. 4A View FIGURE 4 ). Continuous ventral shields present to segment XIV (thoracic chaetiger 12), median ventral groove present from segment XV on (thoracic chaetiger 13) until posterior end ( Figs 4A View FIGURE 4 , 7D,E View FIGURE 7 ).
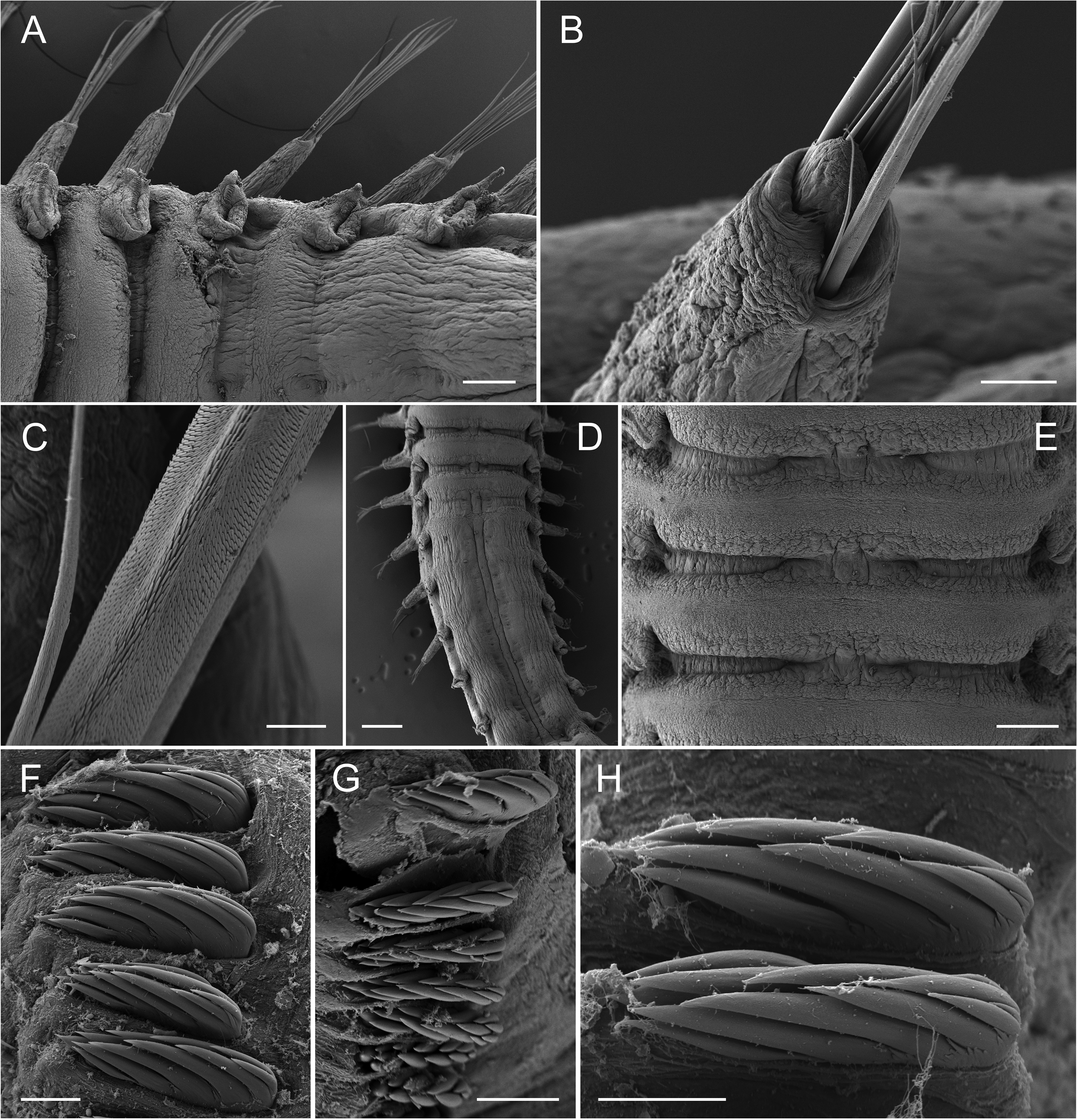
Notopodia with capillary chaetae from segment IV, present in 14 chaetigers; last 12 thoracic chaetigers (starting from segment VI) with neuropodial tori bearing a single row of uncini ( Figs 4A View FIGURE 4 , 7F View FIGURE 7 ); notopodia simple, fingershaped, up to three times longer than wide ( Fig. 7A View FIGURE 7 ); first two notopodia smaller ( Fig. 4A View FIGURE 4 ); tip of notopodia with small terminal lobe (not always visible); notochaetae arranged in semicircle around this lobe ( Figs 4D,E View FIGURE 4 , 7B View FIGURE 7 ). Notochaetae as bilimbate spinulose capillaries, tapering to slender tips; arranged in rows; capillaries from anterior row generally thinner and shorter than those of posterior row ( Figs 4D,E View FIGURE 4 , 7B,C View FIGURE 7 ).
Thoracic neuropodia oval, last three to four neuropodia with short dorsal cirrus ( Figs 4A View FIGURE 4 , 5B View FIGURE 5 , 7A View FIGURE 7 ), anteriorly with small dorsal papilla (at least on two thoracic uncinigers, sometimes on all) which grows into a cirrus that becomes longer towards the posterior end of the thorax ( Fig. 7A View FIGURE 7 ); cirrus length about the same size as the base of respective neuropodium ( Fig. 7A View FIGURE 7 ). Thoracic uncini with five to six teeth in two vertical rows above rostral tooth ( Figs 5D View FIGURE 5 , 7F View FIGURE 7 ); 46–52 uncini on one torus for specimens between 13–16 mm length (largest specimen with 55, and smallest specimen with 34 uncini). Two intermediate segments with neuropodia with very long dorsal cirrus; cirrus usually 2 times longer than base of the respective neuropodium (varies from the same length to 3 times longer); notopodia absent; uncini resemble thoracic uncini ( Figs 5D View FIGURE 5 , 7G View FIGURE 7 ); 39–44 for specimens between 13–16 mm length (largest specimen with 43, and smallest specimen with 27 uncini). Notopodia on abdomen absent. Ten abdominal neuropodia formed as pinnules with very long dorsal cirrus; the ratio of the length of the cirrus to the length of the base of the respective pinnule is 1–3.5:1 ( Figs 5C View FIGURE 5 , 6G View FIGURE 6 ); pinnules of different shapes, rounded, two- or three-lobed ( Fig. 5C View FIGURE 5 ); last six abdominal pinnules can be elongated, and last abdominal pinnule always elongated ( Fig. 5C View FIGURE 5 ). Abdominal uncini with four to five teeth in two vertical rows above rostral tooth ( Figs 5E View FIGURE 5 , 7H View FIGURE 7 ); 47–56 uncini on one pinnule for specimens between 13–16 mm length (largest specimen with 67, and smallest specimen with 35 uncini). Well-developed papillae (glandular pads?) above tori of intermediate segments and above first one to four pinnules of abdominal segments ( Fig. 5B View FIGURE 5 ). Pygidium with terminal anus; 7–17 anal cirri of different lengths and thicknesses, arranged in two more or less distinct rows ( Figs 4B View FIGURE 4 , 6F,G View FIGURE 6 ).
Methyl blue staining pattern. Intense staining of prostomium, bases of noto- and neuropodia and cirri. Prominent bands on ventral thorax (continuous ventral shields), less intensely stained towards the end of thorax, not stained in abdomen. Papillae (glandular pads?) above neuropodia on intermediate and abdominal segments and branchiae stained. Nephridial papillae only visible immediately after intense staining. Rest of body evenly spotted blue, sometimes lighter band laterally between noto- and neuropodia ( Fig. 4A,B,D View FIGURE 4 ).
ShirlastainA staining pattern. Staining pattern equivalent to methyl blue staining, but one additional structure visible: small lobe between paleae-bearing chaetiger (segment II) and first thoracic notopodium (segment IV), most likely vestigial notopodium of segment III ( Fig. 4C View FIGURE 4 ).
Remarks. One thoracic segment was missing in one of 15 specimens, resulting in 14 instead of 15 thoracic chaetigers and 11 instead of 12 thoracic uncinigers. We recommend always examining both sides of specimens, as number of chaetigers and uncinigers are important characters for distinguishing taxa ( Reuscher et al. 2009). One specimen was considerably smaller (7.5 mm long) and thinner (1.1 mm wide), but last six segments were missing. Branchiae were contracted in three specimens and therefore, noticeably shorter and thicker; distance between branchiae for those specimens was only one or even less than one times shorter than branchial bases. An important character of A. cirrata is the presence of a very long dorsal cirri on the neuropodia of the intermediate segments ( Fig. 5B View FIGURE 5 ).
Ampharete cirrata has papillae-like elevations on the intermediate and the first abdominal segments. Imajima et al. (2012) called these structures ‘glandular pads’. Parapar et al. (2012) showed for A. finmarchica that these structures can also be ciliary tufts. The specific structure in the different Ampharete species and their possible homology needs to be clarified urgently.
Uncini of the abdomen are smaller than uncini of intermediate and thoracic segments. However, uncini vary in size along one neuropodium, larger uncini were found at dorsal side of the neuropodium, continuously getting smaller towards ventral side. Extracted uncini possibly originated from different zones of the neuropodia and therefore, size differences between thoracic and intermediate as well as abdominal uncini might be less pronounced. As above described, number of uncini on one neuropodium differed depending on the total body size of specimens. More specifically, the larger the specimen the more uncini were present on one neuropodium. The pygidium showed variability regarding number, length and thickness of anal cirri. It can be difficult to describe the pygidium, as it is often contracted in fixed specimen, sometimes inside the posterior end. One specimen had longer cirri dorsally and shorter and thinner cirri ventrally. A yellow-brown pigment band ventrally on thoracic chaetiger 5 was observed in almost half of the specimens collected between 2002–2010 (fixed in formaldehyde solution and preserved in EtOH), but no pigmentation was found in fresh material from 2019 (treated as described above).
All valid 45 species of Ampharete and 10 synonyms were examined regarding the presence of the combination of the following three characters: thin long paleae, long anal cirri (rather than anal papillae) and neuropodial cirri. Of these species only two, previously synonymized with A. acutifrons , possibly exhibit this combination of characters, A. intermedia and A. cirrata . The original description of Amphicteis intermedia Marion, 1875 was brief, describing the presence of sharp and long paleae, but not mentioning any of the other characters. Since there are no other records of this species and it was not possible to examine it, we decided to disregard A. intermedia . This species needs further investigations (species inquirenda). The description of Ampharete cirrata Webster & Benedict, 1887 from the Northwest Atlantic (Maine, USA) was also short, but contained drawings of the neuropodia. Despite all our efforts, we could not obtain the syntypes of A. cirrata from the NMNH in Washington. However, neither the description nor the photographs ( Fig. 8 View FIGURE 8 ) that we obtained from the NMNH indicated significant differences between the Baltic Sea species and A. cirrata . The specimens found in the Baltic Sea are therefore assigned to A. cirrata . Ampharete cirrata is formally deleted here from the synonym list of A. acutifrons and should be considered a valid species.
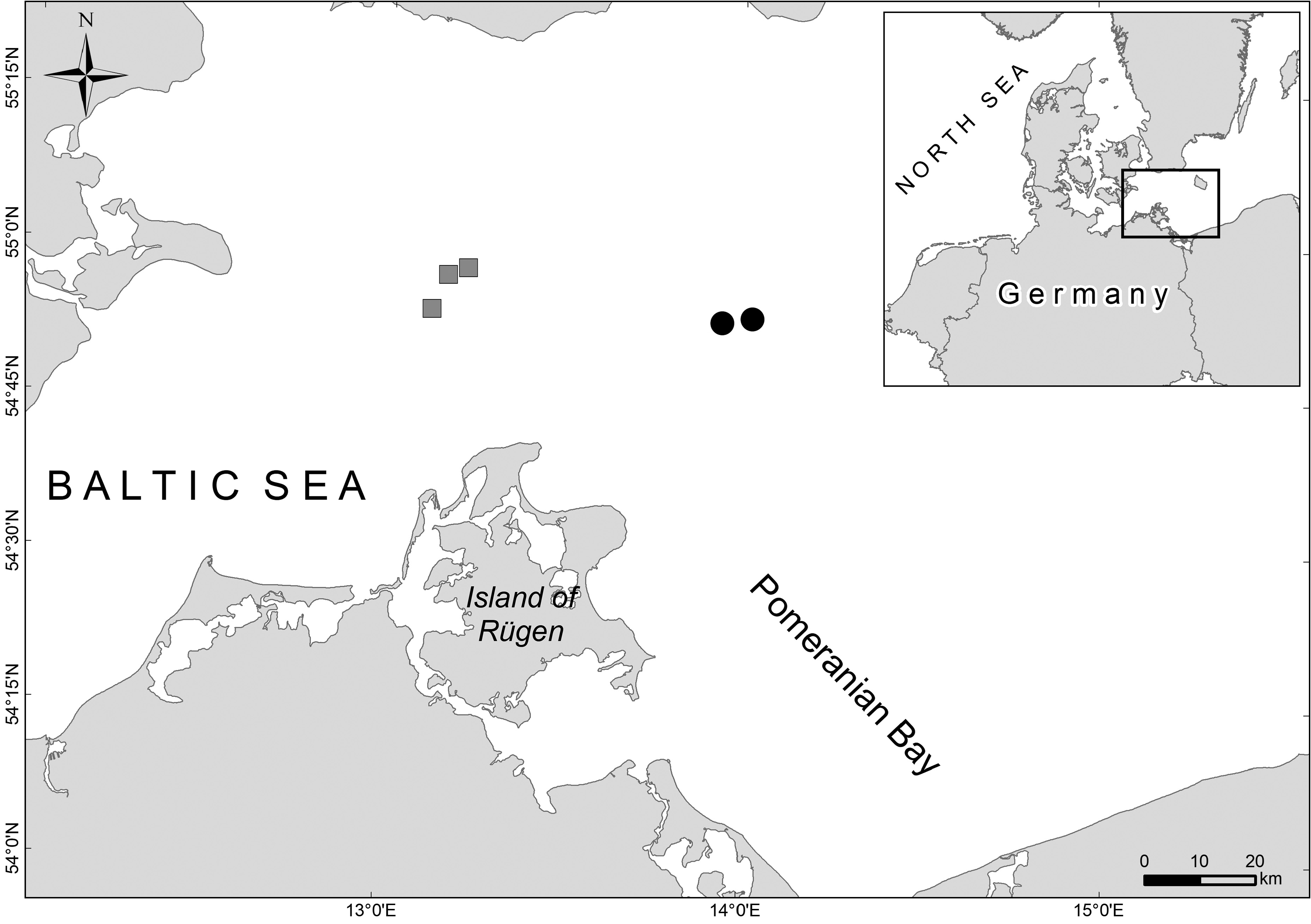
Distribution and ecology. Ampharete cirrata was originally described from the Northwest Atlantic (Maine, USA). Molecular investigations suggest that the distribution of A. cirrata extends beyond the Northwest Atlantic into the Northeast Atlantic (see genetics). The studied material of Ampharete from the Baltic Sea, now assigned to A. cirrata , originates from an area about 30 km north of the island of Rügen ( Fig. 1 View FIGURE 1 ). Specimens were collected in silty sediments of moderate salinity (10–24 psu) in depths of 40–50 m ( Table 1 View TABLE 1 ). Further unpublished records indicate that A. cirrata is also present in shallower localities of about 16 m. The species densities reached from 10–180 individuals/m 2. Oval eggs of about 100–150 µm in diameter were observed in segments XI-XXII in several specimens collected in October.
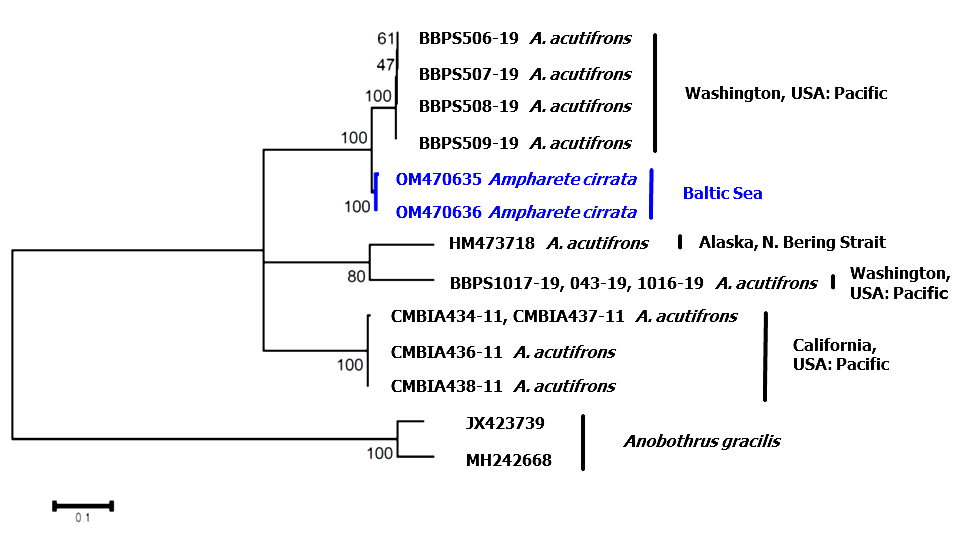
Genetics. In total, 1116 base pairs of the mitochondrial [16S (n = 3) + COI (n = 9)] and 3831 base pairs of the nuclear [18S rDNA (n = 3) + 28S rDNA (n = 1) + Histone H3 (identical to DQ779715 View Materials , see below; n = 9)] of A. cirrata were sequenced. GenBank accession numbers of all obtained sequences are given in Table 3 View TABLE 3 . There were no differences between individuals for the three nuclear genes. For COI and 16S, two haplotypes each were detected, differing 0.2% and 0.3% from each other, respectively. Phylogenetic analysis for the mitochondrial COI fragment included newly generated sequences of A. cirrata and all alleged A. acutifrons sequences from GenBank and BOLD. Two sequences of Anobothrus gracilis were used to root the tree. Excluding the specimens from the Baltic Sea, we found four distinct clades within the alleged A. acutifrons species ( Fig. 9 View FIGURE 9 ). The analysis revealed an A. acutifrons species from Washington, USA to be most closely related to A. cirrata with a divergence of about 4.5% ( Table 4 View TABLE 4 ). Other species summarized under the name of A. acutifrons differed 13.8–15.3% from A. cirrata ( Table 4 View TABLE 4 ).
After a corresponding alignment, obtained sequences of 16S (one of two), 28S and Histone H3 of A. cirrata from the Baltic Sea were completely identical to the sequences of one specimen from Iceland, which was described as A. acutifrons (GenBank accession numbers: 28S: DQ779673 View Materials , H3A: DQ779715 View Materials , 16S: DQ779601 View Materials ( Rousset et al. 2007)).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ampharete cirrata Webster & Benedict, 1887
| Krüger, Lotte, Dietrich, Anna, Bastrop, Ralf & Bick, Andreas 2022 |
Ampharete cirrata
| Webster, H. E. & Benedict, J. E. 1887: 748 |