
Amiracarus pliocennatus Miko, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3670.4.8 |
|
publication LSID |
lsid:zoobank.org:pub:F2EB5077-CE3C-46B3-9E6D-0A4FE4B99BD7 |
|
persistent identifier |
https://treatment.plazi.org/id/2D02E41C-3906-8D2C-E8D9-0261FD9DFDC2 |
|
treatment provided by |
Felipe |
|
scientific name |
Amiracarus pliocennatus Miko |
| status |
|
Description of Amiracarus pliocennatus Miko n. sp.
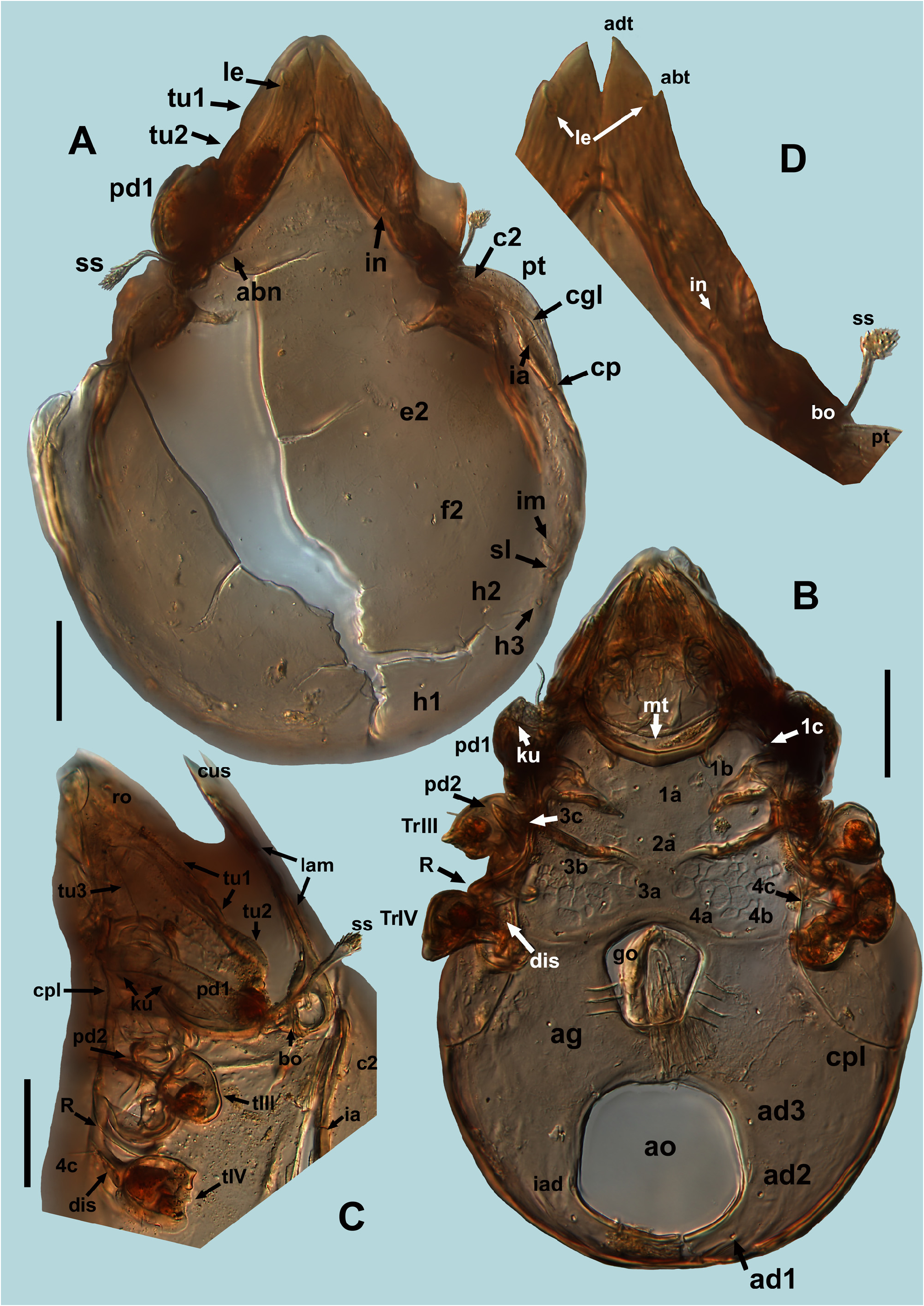
Material examined. Three individuals were found in fossil deposits of Slovenian caves (see the detailed information in Material), one from Late Pliocene sediments of Trhlovca Cave , and other two from the Early Pleistocene in Račiška pečina Cave. The best preserved individual from Trhlovca Cave ( Fig. 6 A View FIGURE 6 ) is designated as holotype, the other individual ( Fig. 6 C View FIGURE 6 ) as paratype. Holotype was mostly used for description of all characters, paratype provided for more information on measurements, lateral and ventral characters. Third individual ( Fig. 6 B View FIGURE 6 ) was lost after drawing some, mostly ventral, characters. Holotype will be deposited in acarological collection of the Czech National Museum in Prague, paratype will be deposited in the Collection of first author .
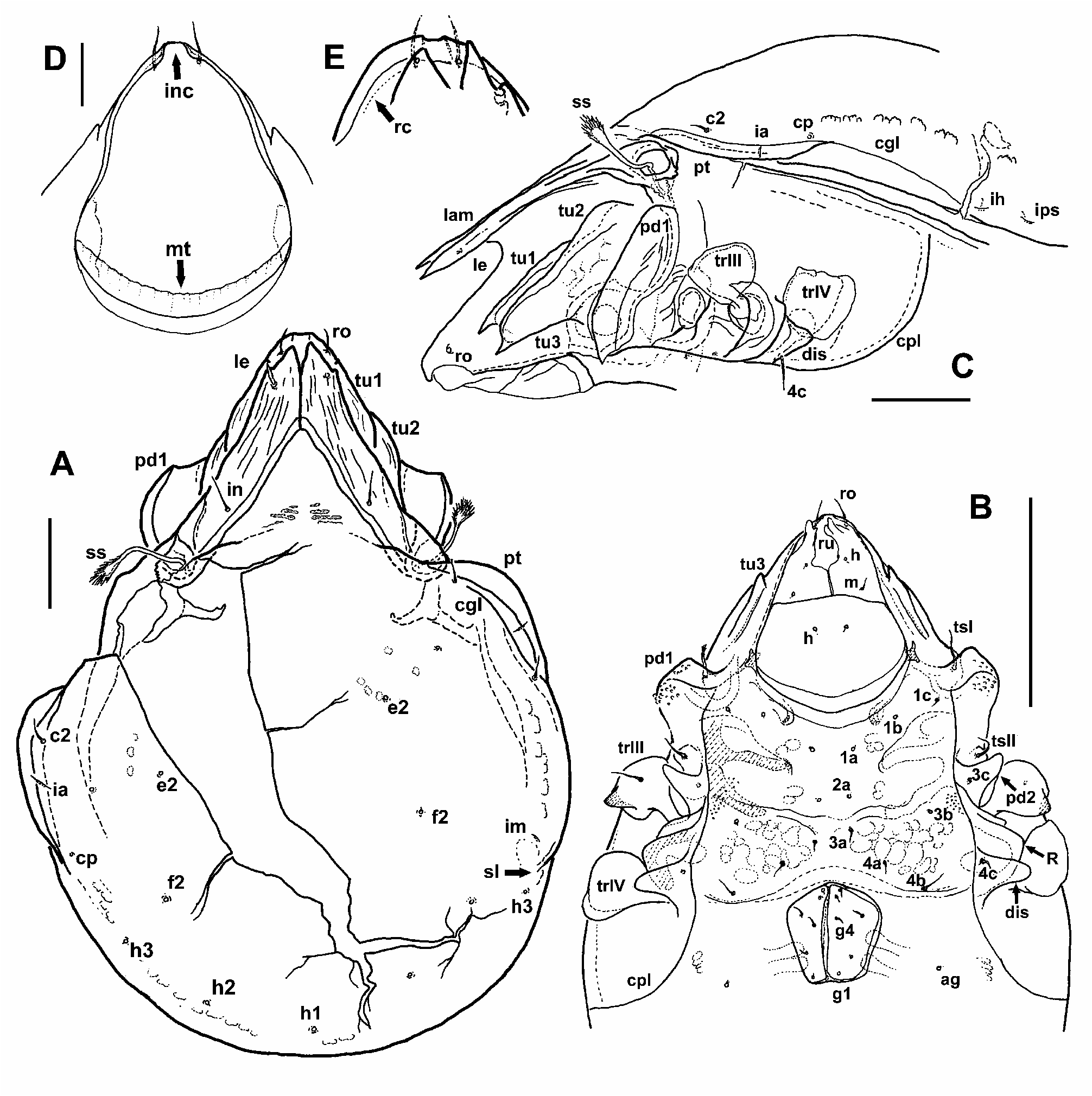
Diagnosis. Large Amiracarus with relatively long interlamellar seta (17–18 µm), inserted on the lamella close to their inner margin. Rostrum dorsally with shallow incision, rostral seta inserted at the end of short carina (rc) running parallel with lateral margin of rostrum ( Fig. 3 E View FIGURE 3 ). Lamella large but not completely covering the rostral part of prodorsum, adaxial margins of lamellar cusps proximally parallel, but clearly diverging in distal part (in last quarter of the cuspis length). Large adaxial tooth of cuspis always developed, more or less sharp, small abaxial tooth either present or strongly reduced to a small tubercle, or even completely absent. Insertion of lamellar seta sub-apical or positioned in cusp incision, closer to small abaxial tooth. Tutorium complex, with two acute cusps anteriorly as in other species of the genus. Pteromorph developed as narrow rounded blade, notogastral setae smooth, relatively long (7–13 µm). Setae e 2 and f 2 inserted in the middle of the notogaster, not shifted posteriad; e 2 positioned at about the level of seta c p or even slightly more anteriad, seta h 3 positioned posteriad to lyrifissure im and lateromedial furrow. Trochanters III and IV ventrally with bladelike auricular carina, projecting distally as rounded but distinct spur. Ovipositor with long distal setae on its lobes.
Description of adult (immatures unknown)( Figs 3–4 View FIGURE 3 View FIGURE 4 , 6–9 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ).
Measurements. Precise measurements difficult because of the poor state of available individuals, the values should be therefore seen as best possible approximation. Total body length 332–345 µm, length of prodorsum 87– 97 µm, length of notogaster 235–258 µm, ratio of prodorsum and notogaster length from 1:2,4 to 1:3. Maximum width of prodorsum 148–166 µm, maximum width of body (notogaster) 228–262 µm (n=3 for all measurements). Length of lamellae 95–120 µm, length of lamellar cuspis 40–48 µm.
General characters. Body form as usual for Amiracarus , relatively broad. Colour yellowish to reddish brown. Without visible stronger layers of cerotegument dorsally, granular cerotegument observed laterally on pedotectum I. Lamellae and partly also tutorial costulae with longitudinal fine striation, appearing rugose. Some individuals with partly macerated cuticle and consequently with net-like pattern, more transparent and much lighter coloured ( Fig. 6 View FIGURE 6 ). Body of found fossil individuals damaged on many places and covered by artefacts, making observation of morphological details difficult.
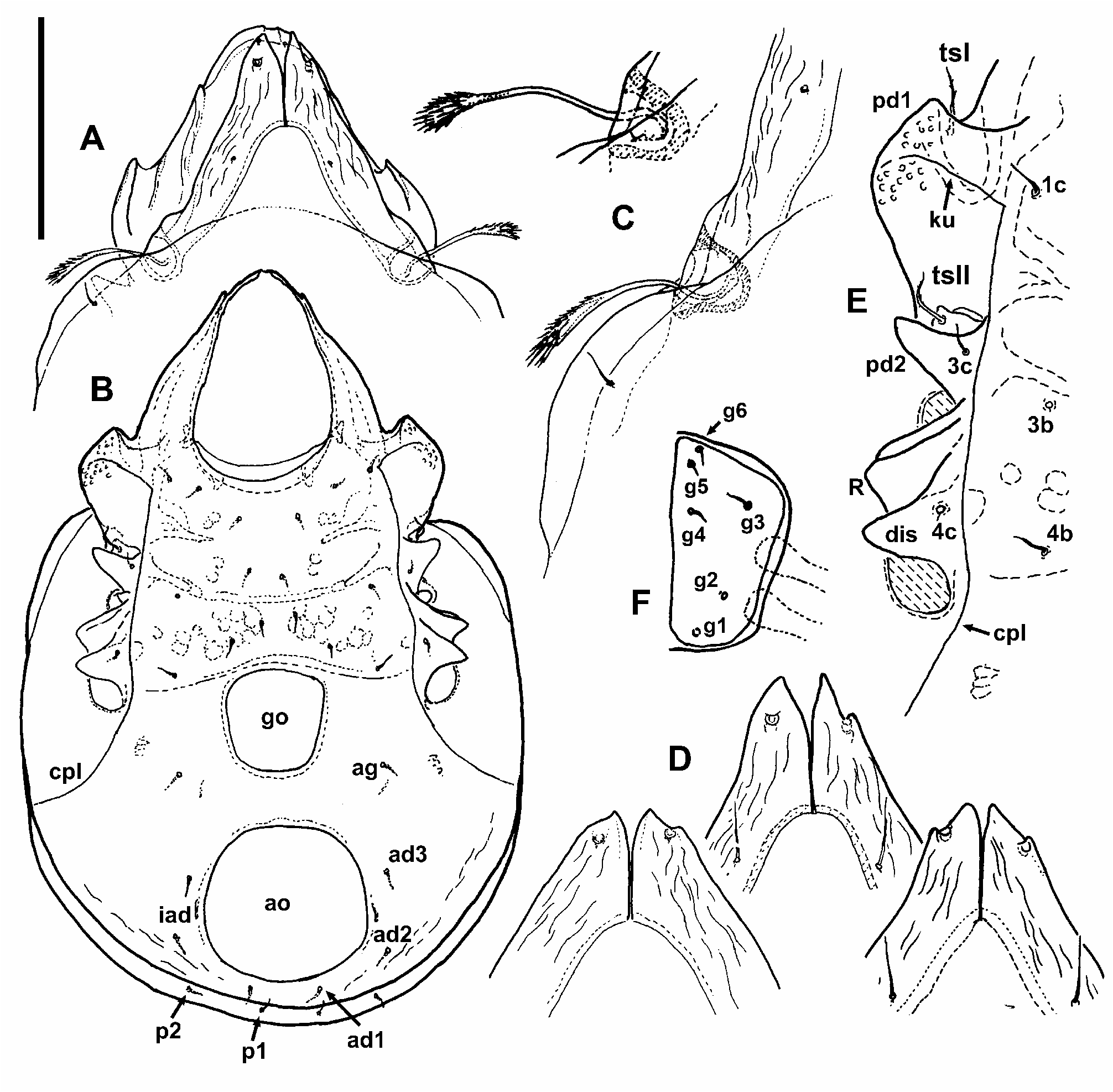
Prodorsum. Broad and relatively short in shape. Rostrum broad, not fully covered by lamellae, appearing slightly angulated in dorsal view. Rostral incision appearing relatively shallow in dorsal view but in fact very deep and distinct when seen ventrally or anteroventrally ( Fig. 3 D View FIGURE 3 ). Insertion of rostral seta positioned close to the margin of the rostrum, in one of the studied individuals with better preserved rostral part was the insertion at the end of short rostral carina, running from insertion lateroposteriad towards tutorial tip ( Fig. 3 E View FIGURE 3 ). This carina was hardly observable in other individuals. Rostral seta short (about 14 µm), smooth and fine, slightly bent. Lamellae well developed and broad, converging anteriomediad, more or less of the same width throughout, covering large part of the prodorsum. Lamellar cusps distinct, long and broad, their adaxial margins in proximal three quarters parallel, only distally diverging ( Figs 4 D View FIGURE 4 , 8 A–B View FIGURE 8 ). Adaxial tooth of cuspis strong and large, more or less sharp. Abaxial tooth or blunt tubercle at its position also usually present, in some individuals unilaterally or completely absent, making shape of lamellar cuspis variable ( Fig. 4 D View FIGURE 4 ). Insertion of lamellar seta often sub-apical, on the dorsal surface of cuspis at a distance from the margin, or positioned on straight or shallowly incised lateral edge of cuspis between the teeth, closer to abaxial one. Bothridium large, in dorsal view hidden (sunken) between base of lamella and another subbothridial lamella, ( Fig. 7 A, C View FIGURE 7 ). Sensillus generally resembling the shape and size known from other species of the genus, still rather variable ( Figs 4 C View FIGURE 4 , 8 D–E View FIGURE 8 ) either with long and flexible stalk and short, sub-spherical or club-shaped head, or head more elongated with a much shorter stalk. Head of sensillus always covered with dense barbs, relatively long and dark coloured. Interlamellar seta smooth and fine, relatively long (about 18 µm), as long as, or even slightly longer than width of proximal part of lamella ( Fig. 3 A View FIGURE 3 ), inserted on the lamella, closer to its adaxial margin. Interlamellar area triangular or arched anteriorly, in the middle with three more or less distinct pairs of muscular sigillae, anterior ones being the smallest. Tutorium, seen in lateral view, developed similarly as in other species of the genus ( Figs 3 C View FIGURE 3 , 8 F–G View FIGURE 8 ). Tutorial carinae 1 and 2 well developed, striated, visible partly also in dorsal view, tutorial carina 2 appearing longer and slightly S-shaped. Anterior end of each of the tutorial carinae 1 and 3 with distinct and sharp projection (cusp). Pedotectum I well developed and large, its anterior margin not reaching beyond half of the prodorsum length in dorsal view.
Notogaster. Relatively broad in shape, with indistinct anterior border between the lamellae. Row of muscle sigillae well visible around the lateral and posterior margin, two groups of sigillae also present anteriorly on notogaster, more or less behind bothridia and proximal part of lamellae. Lateromedial furrow (sl) present, even if not always apparent in dorsal view. Pteromorph developed as rounded, narrow blade ( Figs 3 A, C View FIGURE 3 ; 9 B View FIGURE 9 ). Circumgastric line clearly visible throughout most of the pteromorph length, almost reaching anterior margin of notogaster. Lyrifissure ia closer to insertion of seta c p than to c 2, lyrifissure im anterior to seta h 3. Notogastral setae mostly lost in studied individuals, only insertions observable, setae c 2 and c p preserved, moderately long (12–13 µm), smooth and fine, similarly as in known extant species of the genus. Setae e 2 and f 2 inserted more anterior than usual, e 2 at the level or even anterior to level of setae c p, position of other setae similar to other known species of the genus.
Gnathosoma. In general similar to extant species. Camerostome broadening posteriorly, prolonged and narrowed anteriorly, appearing piriform. Incision of rostral hood well visible anteriorly. Mentotectum wide and slightly rugged ( Fig. 3D View FIGURE 3 ), with slightly undulated anterior edge. Infracapitulum ( Figs 3B View FIGURE 3 , 9 A View FIGURE 9 ) diarthric, developed similarly to that in extant species, with rutella partly modified and forming rutellar microtube (see Wauthy &Ducarme 2011). Setae a, m and h relatively short, fine and smooth. Other characters of mouthparts difficult to observe in studied individuals or invisible (partly also lost).
Ventral characters. Generally as in other species of the genus. Epimeral region and ventral plate laterally surrounded by circumpedal line, which is distinct, well developed, reaching anteriorly carinae ku on pedotectum I. Lateral parts of podosoma with usual structure ( Fig. 4 E View FIGURE 4 ), pedotectum I large but appearing relatively short anteriorly, pedotectum II narrow and acute. Discidium large, perpendicular to body axis. Acetabular protrusion R large, with distinct ventral carina kt. Epimeral area with muscular sigillae partly forming a net-like structure, more apparent on epimeres III and IV. Apodeme II almost transversal, relatively long, sejugal apodeme shorter, oblique, with end pointing more or less to anterior part of genital opening. Epimeral setal formula 3–1–3–3, with setae 3c and 4c positioned laterally from the circumpedal line, on pedotectum II and discidium, respectively ( Fig. 4 B, E View FIGURE 4 ). Posterior margin of epimeres IV with transversal band, almost straight in lateral parts, curving slightly anteriad anterior to genital plates ( Figs 3 B View FIGURE 3 , 4 B View FIGURE 4 ). Anogenital field normally developed, rugged posteriorly (appearing as indistinctly striated), anal aperture larger than genital, broadly oval to subcircular ( Fig. 4 B View FIGURE 4 ). Only two pairs of genital papillae observable in ventral view. Ovipositor (op, Fig. 9 D View FIGURE 9 ) relatively short, with very long distal setae on its lobes (τ 1 and ψ 1 about 18 µm, other setae on lobes about half of this length), and setae k smooth and short. Genital plates with 6 smooth, fine and short setae each, anal plates not preserved in our material. Positions and numbers of aggenital and adanal setae as usual in genus, i.e. 1 pair of aggenital and 3 pairs of adanal setae present, lyrifissure iad behind the insertion of ad3, paraanal.
Legs. Almost completely lost in studied individuals, only trochanters present. Trochanters I and II with rather long setae, well visible in ventral view, trochanteral seta I may bear small barbs, trochanteral seta II smooth ( Fig. 4 E View FIGURE 4 ). Trochanter III with distinct ventral spur ending by distinct but blunt tip. Trochanter IV with similar projection, which is broadly rounded, laminar, without tip ( Fig. 4 E–F View FIGURE 4 ). Smooth ventral seta on trochanter III preserved, relatively long.
Derivatio nominis. The specific name pliocennatus indicates presence of the species in Pliocene (and Pleistocene) periods: pliocen(e) -natus: born in Pliocene.
Remarks. Some of the characters found on the fossil material of Amiracarus pliocennatus Miko , n. sp. seem to be rather variable (lamellar cuspis, form of sensillar head). Same variability was observed also within the same individual (left and right cuspis of lamellae, left and right sensillus), so both specimens found together in the same fossil layer were attributed to the same species. Observed slight differences of holotype, although from another cave, were in our view within the same range of accepted variability, so all found individuals are considered to belong to the same species. The new (fossil) species is similar to extant ones, but can be distinguished by overall larger size, development of lamellae with a tendency of loosing abaxial tooth, notogastral setation with setae e 2 and f 2 positioned unusually more anteriad than in other species and by other less apparent characters. Also setation of ovipositor seems to be particular, with setae on ovipositor lobes much longer than those from extant species, as described by Wauthy &Ducarme (2011) and Lions (1978). Some characters of all known species of the genus are summarised in Table 1.
The relationships of known extant species remain in some aspects unclear. Wauthy &Ducarme (2011) provided an extremely detailed description of A. grootaerti , species found in Belgian caves (described as Miracarus ), including differences from A. senensis and, in more detail, from A. abeloosi . While the differences between grootaerti and abeloosi are clear, and both these species could be distinguished easily by macro-characters (the most remarkable is the different development of lamellae and lamellar cuspis and anterior margin of notogaster), the differences between grootaerti and senensis appear to be much finer and more questionable. The major distinguishing character, used by Wauthy & Ducarme, is the absence of the rostral incision in senensis . Nevertheless, based on figures (Figs. Ia, Ib and IIc) provided by Bernini (1975), the presence of a slit is indirectly indicated, as the rostral part seems to be developed very similar to the one present in our fossil material if seen dorsally or dorso-laterally, and a rostral incision is usually only visible in ventral or anterior view. Therefore, the option that a slit was just overlooked or not mentioned by Bernini in his description cannot be excluded, and confirmation of its absence is needed. When comparing a whole set of other characters, Wauthy &Ducarme (2011) indicate identical development in both grootaerti and senensis , with a single exception relating to leg setation (proral and primiventral setae of tarsi). Therefore, if the absence of rostral slit is not confirmed in senensis , and other characters not mentioned by Bernini (such as very fine, but visible margin of anterior notogastral tectum) are found in Italian species, further distinguishing characters will be necessary—otherwise there is a question of potential synonymy. Our fossil material from Romania (see above) has been attributed to senensis , based on overall similarity, body proportions, ventral characters, and also anterior margin and setation of notogaster, but a rostral slit was observed here (visible in ventral view). In any case, both species are very closely related even if synonymy is not confirmed. Another similar problem is related to the taxa abeloosi and similis . Subìas in his “Listado sistemático (2004, internet version 2012) considers these taxa to be synonyms. Based on their descriptions, however, they differ clearly by body length, and also by development of lateral parts of the podosoma, namely acetabular protrusions R and discidia. Obviously, this depends on the accuracy of figures in descriptions, and again needs to be checked on original material. Even here, synonymy is not completely excluded. For the time being, using the above mentioned criteria, all taxa are considered as separate, valid species until possible synonymies are confirmed. All known species of Amiracarus can be distinguished by the following key (see also table 1 for specific characters):
1 Adaxial margins of lamellar cusps divergent in more than half of their length...................................... 2
- Adaxial margins of lamellar cusps fully parallel or parallel in more than half of their length.......................... 4
2 (1) Interlamellar seta inserted very close to inner margin of lamella and close to anterior margin of notogaster. Notogastral seta h 3 shifted forwards, inserted anterior to the lyrifissure im. Discidium very large. Adanal seta ad 3 inserted close to lyrifissure iad near anterior margin of anal opening, distance of insertions ad 2 -ad 3 much larger (2–3 times) than distance ad 1 -ad 2. Body size 284–296 µm................................................................. A. discrepans ( Mahunka 1966) View in CoL
- Interlamellar seta inserted more distant from both inner margin of lamella and anterior margin of notogaster. Notogastral seta h 3 inserted behind the level of lyrifissure im. Distance of insertions ad 2 -ad 3 usually subequal or only slightly larger (less than 2 times) than distance ad 1 -ad 2 ............................................................................. 3
3 (2) Anterior margin of notogaster with tectum, extending over prodorsum and covering almost entirely central part of prodorsum up to the base of lamellar cusps. Anterior margin of this tectum faint but visible. Rostral hood with distinct and deep incision. Notogastral seta f 2 at about the level of seta h 3, forming one more or less transversal row. Body size 313–334 µm................................................................................ A. grootaerti ( Wauthy &Ducarme 2011) View in CoL
- Anterior margin of notogaster axially indistinct, tectum not observable or visible only as a short line in vicinity of interlamellar setae, not entirely covering central part of prodorsum. Rostral incision not visible in dorsal view (see remarks above). Notogastral seta f 2 behind the level of seta h 3, almost at the level of h 2 or even behind. Body size 295–320 µm.................................................................................................. A. senensis ( Bernini 1975) View in CoL
4 (2) Lamellar cuspis long, covering the rostrum, reaching or even overreaching its anterior margin in dorsal view. Teeth of cuspis medium to small, sub-equal in size, their tips reaching anteriad about the same distance. Incision between teeth of cuspis shallow and short, lamellar seta inserted about in the middle between them. Anterior margin of notogaster distinct, well visible.. .................................................................................................... 5
- Lamellar cuspis shorter than the rostrum, thus rostrum at least partly visible in dorsal view. Adaxial tooth of cuspis clearly longer than external, protruding far more anteriad. Abaxial tooth smaller or even fully reduced. Lamellar seta inserted more laterally, closer to external tooth (if present) or on the surface of the cuspis at a distance from anterior margin. Anterior margin of notogaster indistinct in central part. (Only known from fossils from Balkan peninsula.) Body size 332–345 µm.......................................................................................... A. pliocennatus Miko , n. sp.
5 (4) Smaller species (body length 255–275 µm). Acetabular protrusion R smaller than discidium, distance between tips of R therefore significantly smaller than between the tips of discidia.................... A. similis ( Subìas &Iturrondobeitia, 1978) View in CoL
- Larger species (body length 291–318 µm). Acetabular protrusion R developed similarly to discidium, distance between tips of R and tips of discidia subequal....................................................... A. abeloosi ( Lions 1978) View in CoL
Remarks on ecology of the species. While three of the species are known from soil or moss growths ( A. discrepans ( Mahunka 1966) from meadow soil, A. similis (Subìas &Iturrondobeitia 1977) from forest mosses, and part of material of A. abeloosi ( Lions 1978) , collected from beech forest soil), at least part of the species are from caves and their close vicinity ( A. senensis ( Bernini 1975) , A. abeloosi ( Lions 1978) and A. grootaerti ( Wauthy &Ducarme 2011) . This indicates possible affinity of the genus Amiracarus to caves and similar microhabitats. Presence of fossil material in cave sediments is therefore not surprising, and may indicate either broader presence of the genus in habitats surrounding caves, or troglophily/troglobiosis of the species. It is therefore difficult to judge if fossil material of this genus can be used for extrapolations of climatic and ecological conditions in areas around the caves like other species found in sediments. Some Amiracarus species possibly inhabited caves already in the Tertiary and could appear accidentally in sediments together with other oribatid species which were transported into cave from cave environs. In any case, given the relative rarity of all species in extant fauna, their presence in caves and findings of very close relatives in fossil cave sediments strongly suggests the potential relic character of the genus. This may be supported also by the fact of (to date) disjunctive presence over a large area of the Mediterranean and south-west Europe.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |