
Inocelliidae Navás, 1913
|
publication ID |
https://doi.org/10.11646/zootaxa.3784.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:D5E03502-7BD3-41F4-A4CF-5537B1462A23 |
|
DOI |
https://doi.org/10.5281/zenodo.6131171 |
|
persistent identifier |
https://treatment.plazi.org/id/039287A3-FE25-3737-23C7-6C14FD0FFF7A |
|
treatment provided by |
Plazi |
|
scientific name |
Inocelliidae Navás, 1913 |
| status |
|
Family Inocelliidae Navás, 1913 View in CoL View at ENA
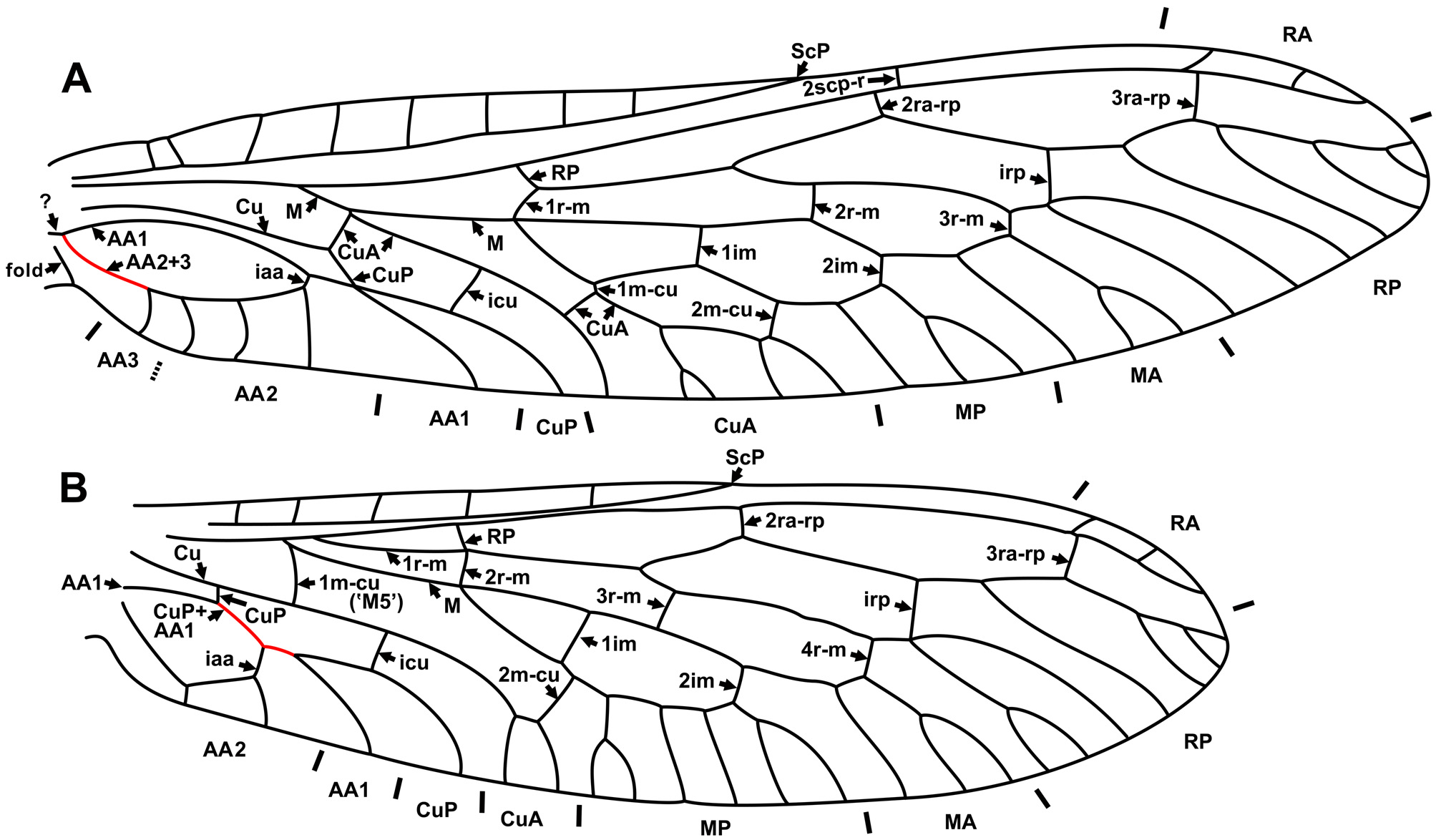
Diagnostic character states of venation. Pterostigma not enclosed by crossvein proximally; incorporated pterostigmal branches of RA absent. Forewing: 2scp-r located near termination of Sc; 3ra-rp always distad pterostigma; three crossveins between RA, RP (few exceptions, below); stem of M+CuA always long (except Sininocellia Yang, 1985 , where relatively short); four crossveins between R/ RP, M/MA; basal crossvein between R, M (1r-m) connect M, R; 4r-m present; in medial space, three crossveins forming three doi; 4doi present, formed by MP, CuA, and two crossveins between MP, CuA (except few species, below); MP, CuA connect by two crossveins (MP, CuA apparently fused as in Raphidiidae in few species with two doi, below); AA1 usually simple (few exceptions, below); rather long free basal portions of AA2, AA3 (before their fusion).
Hind wing: 2scp-r connects ScP and RA; 3ra-rp always distad pterostigma; CuP not fused with AA1.
Composition. Seven extant genera (40 valid species): Inocellia Schneider, 1843 [22 species;Palaearctic region]; Fibla [4 species; Mediterranean]; Parainocellia Aspöck et Aspöck, 1968c [4 species; Europe; Burma, Vietnam]; Sininocellia [2 species; China]; Amurinocellia Aspock et Aspock, 1973 [3 species; East Asia]; Indianoinocellia [2 species; Mexico]; Negha [3 species; North America] ( Liu et al. 2009a, b, 2010b, c, 2012a,b, 2013, 2014; Aspöck et al. 2011, 2012b).
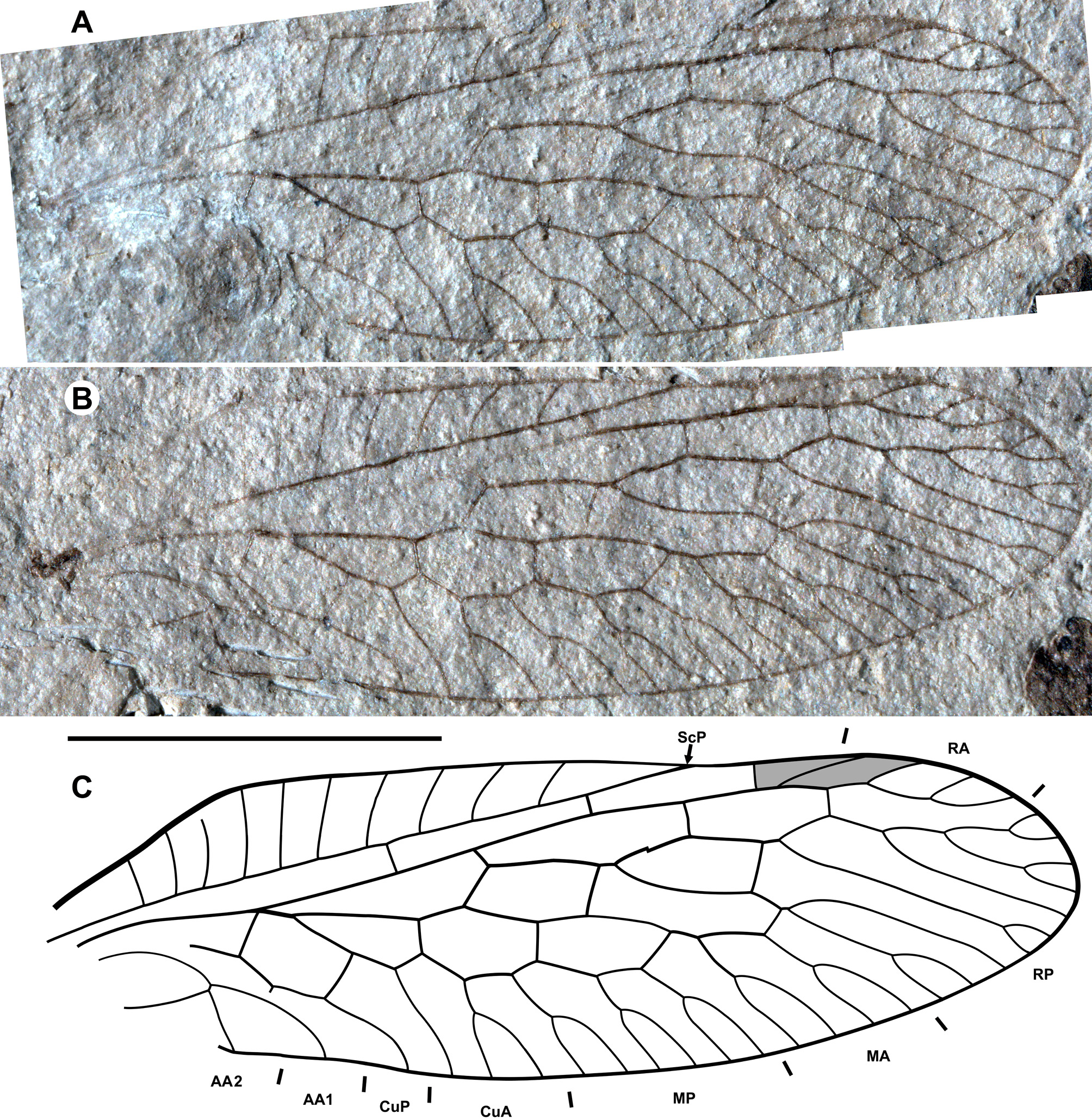
Seven fossil species (six named), all recorded from the Cenozoic: Electrinocellia peculiaris , Succinofibla aperta Aspöck et Aspöck, 2004 , ‘ Fibla ’ erigena (Menge in Pictet-Baraban et Hagen, 1856), ‘ Fibla’ carpenteri Engel, 1998 , and an undesribed specimen ( Janzen 2002: Figs 54, 547) from late Eocene Baltic amber; ‘ Fibla’ exusta ( Cockerell et Custer, 1925) from the late Eocene of Florissant (Colorado); Miofibla cerdanica Nel, 1993 from the late Miocene of Bellver-en-Cerdaña ( Spain) and the unnamed ‘Species G’ from the late Miocene of Murat ( France). Inocelliid larvae are known from Baltic amber ( Weidner 1958; Janzen 2002: Figs 55, 548).
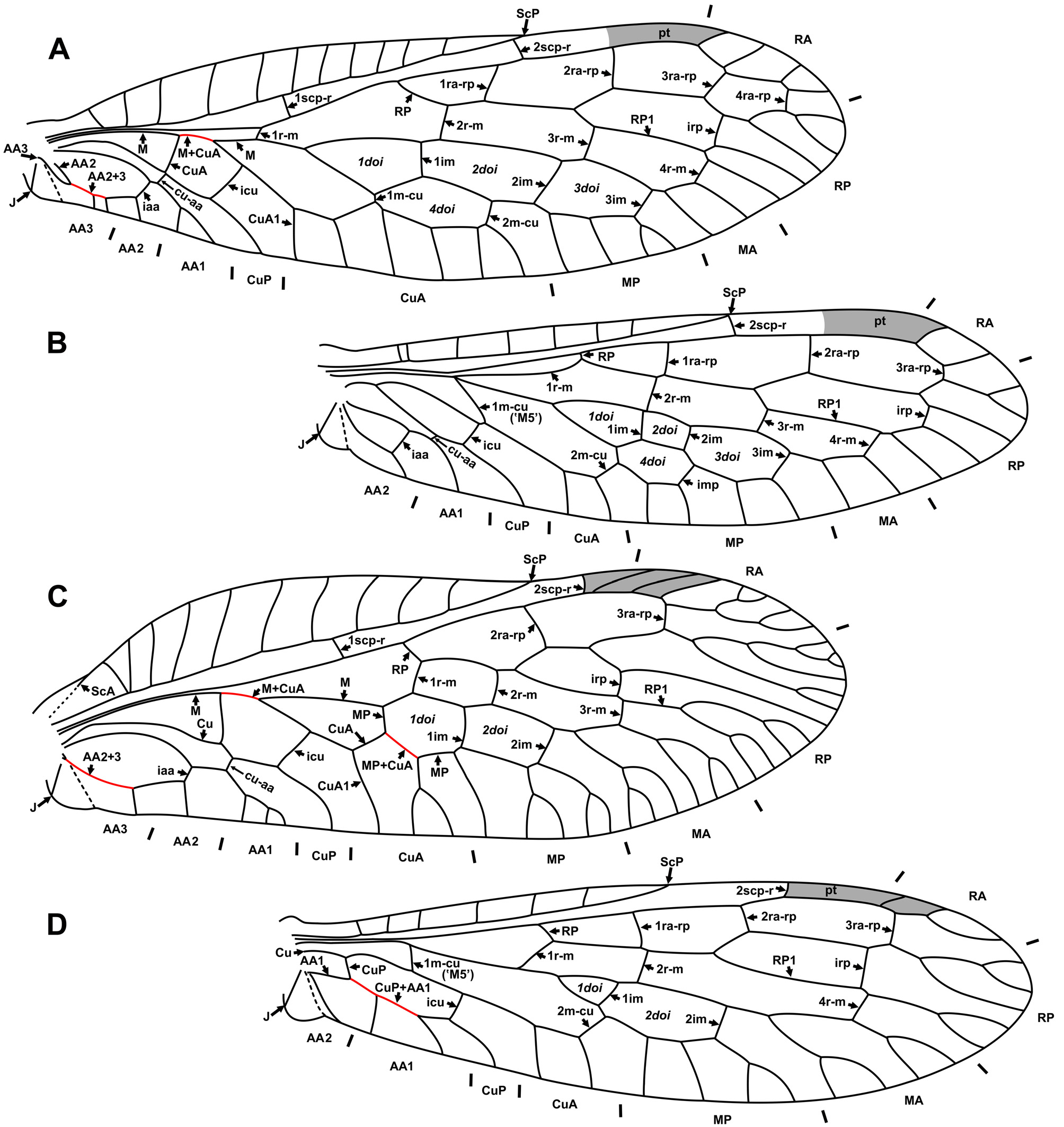
Comments on characters. Radial crossveins. All species of extant Inocelliidae have three radial crossveins between RA and RP in the forewing, with the exception of two species of Sininocellia (four crossveins) and individual specimens of the highly variable Inocellia folvostigmata Aspöck et Aspöck, 1968 (Aspöck & Aspöck 1968: Fig. 1 View FIGURE 1 h) (two crossveins); other specimens of this species have three of these crossveins (e.g., Aspöck et al. 1982: Figs 26–29). Electrinocellia peculiaris from Baltic amber has only two radial crossveins ( Carpenter 1957: Fig. 3 View FIGURE 3 A).
Discoidal cells. Until recently, the vast majority of extant Inocelliidae species were known to have four (3+1) doi in the forewing ( Fig. 2 View FIGURE 2 A). Probably, the single documented exception was three (2+1) doi in the two species Negha meridionalis Aspöck, 1988 ( Aspöck 1988: Fig. 1 View FIGURE 1 ) from California and Inocellia f. fulvostigmata from India ( Aspöck et al. 1982: Figs 26, 27), and two (2+0) doi in Inocellia fulvostigmata nigrostigmata Aspöck et al. 1982 from India (Aspöck & Aspöck 1968: Fig. 1 View FIGURE 1 h; Aspöck et al. 1982: Figs 28, 29). However, the rich inocelliid fauna from eastern and southeastern Asia reported in the last few years shows remarkable venational diversity including three (2+1) doi present in Inocellia biprocessus Liu et al., 2010 from Shandong Province in China, and two doi (2+0) in Inocellia bilobata Aspöck et al., 2011 ; I. cornuta Aspöck et al., 2011 ; Inocellia longispina Aspöck et al., 2011 ; Inocellia striata (all from Thailand, southernmost findings of Raphidioptera in the Old World); and Inocellia yunnanica Liu et al., 2012 from Yunnan Province in China ( Liu et al. 2010c: Fig. 1 View FIGURE 1 ; Aspöck et al. 2011: Figs 1 View FIGURE 1 , 10 View FIGURE 10 , 19 View FIGURE 19 –28, 40–43; Liu et al., 2012a: Figs 3 View FIGURE 3 , 4 View FIGURE 4 ). Electrinocellia peculiaris from late Eocene Baltic amber also has two doi.
Most probably, in these few species of Inocelliidae that have two doi, MP and CuA are fused for some distance similarly to Raphidiidae . At least, the venation of this area in these inocelliid species is not different from that of Raphiididae.
Forewing AA1. The inocelliid AA1 is usually simple. Occasionally, AA1 is forked (often in only one of the forewings) in some extant species, e.g., Sininocellia gigantos Yang, 1985 , S. chikum Liu et al., 2012 (Liu et al. 2012b: Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ), Fibla hesperica Navás, 1915 ( Aspöck et al. 1991: Fig. 45) and Negha inflata ( Hagen, 1861) ( Carpenter 1936: Fig. 7 View FIGURE 7 ); in the fossil Fibla carpenteri , and the unnamed species ‘Espèce G (probablement Fibla )’ ( Nel 1993: Fig. 12 View FIGURE 12 ). It is noteworthy that all of these species (except Negha inflata ) possess a long basal 1r-m crossvein (brace) in the hind wing, a plesiomorphic character state. We suspect that the forked AA1 is a plesiomorphic state in this family as well.
Remarks. All but one fossil species have wings similar to those of the extant genera Fibla and Sininocellia in the presence of the long basal r-m brace (1r-m), a plesiomorphic state in Neuropterida. The exception is Electrinocellia peculiaris . This species has two discoidal cells, two radial and medial crossveins in the forewing (as in raphidiids) and 1r-m in the hind wing is lost (as in most extant inocelliids; this condition is only known in this species among fossil taxa of the family). Grimaldi (2000) believed that “it shares inocelliid and raphidiid features” (p. 262). Engel (1998) separated it into the monotypic inocelliid subfamily Electrinocelliinae. However, some recently described species of Inocellia from southeastern Asia have very similar venation (see Aspöck et al. 2011: Figs 23 View FIGURE 23 –28), discrediting the validity of this subfamily.
Two Mesozoic species have been assigned to the Inocelliidae . The Early Cretaceous Sinoinocellia liaoxiensis Wang, 1987 was attributed to this family ( Wang 1987: Fig. 17 View FIGURE 17 ); however, the drawing of this incompletelypreserved species clearly shows that it belongs to Mesoraphidiidae (s.l.) by its forewing character states, e.g., only two crossveins between RA and RP (three in Inocelliidae ); three crossveins between R/ RP and M/MA (four in Inocelliidae ); 1r-m connects R with MA, not M as in all Inocelliidae . Also, an undescribed species from the Late Cretaceous (Turonian) locality of Kzyl-Dzhar (now Kyzylzhar) was thought as belonging to Inocelliidae ( Zherikhin 1978, p. 78). This specimen also belongs with certainty to Mesoraphidiidae (s.l.); it is most similar to Alloraphidia dorfi Carpenter, 1968 (VM, pers. obs.).
No venational autapomorphies of Inocelliidae were identified by Aspöck and Aspöck (2004). However, we find two forewing character states to be autapomorphies of this family: (1) the presence of the crossvein 1ra-rp, and (2) the crossvein 1r-m connects M and R (see above).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |