
Aega Leach, 1815
|
publication ID |
https://doi.org/ 10.5281/zenodo.5391872 |
|
publication LSID |
lsid:zoobank.org:pub:67B610D8-3A59-4571-9D65-031EAB044EC3 |
|
persistent identifier |
https://treatment.plazi.org/id/EF52F314-FFEA-695A-3305-FB86FD30F945 |
|
treatment provided by |
Marcus |
|
scientific name |
Aega Leach, 1815 |
| status |
|
Genre Aega Leach, 1815 View in CoL
Le genre Aega , établi par Leach (1815), a d’abord été insuffisamment caractérisé en particulier par Schioedte & Meinert (1879). Par la suite, des diagnoses en ont été données successivement par Brusca (1983) et Kensley & Schotte (1989). À cette occasion, Brusca (1983) a divisé le genre Aega en deux sous-genres, Aega et Rhamphion, entre lesquels l’auteur a réparti toutes les espèces décrites jusqu’alors. Par la suite, ces deux sousgenres n’ont toutefois pas été acceptés par tous. Kensley & Schotte (1989) ont retenu cette distinction pour les espèces Aega (Aega) deshaysiana ( Milne Edwards, 1840) , A. (Aega) ecarinata Richardson, 1898 , A. (Rhamphion) dentata Schioedte & Meinert, 1879 et A. (Rhamphion) tenuipes Schioedte & Meinert, 1879 , des Antilles. Wetzer (1990) a également décrit un Aega (Rhamphion) francoisae , récolté dans le cloaque d’une ascidie au niveau des Îles Galapagos. Par contre, Bruce (1988) n’a pas retenu la proposition de Brusca pour la nouvelle espèce Aega leptonica de Floride. Mais, c’est surtout en 1996, à l’occasion de la description de l’espèce Aega komai du Japon, que Bruce (1996) a fait observer que certaines espèces ne s’adaptaient pas exactement à une telle définition de deux sous-genres et a donc préféré conserver le genre Aega . Ces conclusions de Bruce (1996) ont par la suite été retenues par Kensley & Chan (2001) dans leur description de la nouvelle espèce Aega falcata de Taiwan. Plus récemment, Bruce (comm. pers.) a proposé de ne pas utiliser, au moins momentanément et en attendant des recherches plus précises, les deux sous-genres tels que définis par Brusca (1983).
Compte tenu de l’état actuel de nos connaissances sur ce sujet, pour les trois espèces de Nouvelle-Calédonie, nous conservons donc le genre Aega . On peut toutefois souligner que, selon Brusca (1983), ces trois espèces appartiendraient au sous-genre Aega (Aega) .
Précisons également que, à partir des seuls spécimens dont nous disposions, il ne nous a pas été possible d’observer et donc de représenter, tout au moins avec suffisamment d’exactitude, l’ensemble des cinq articles du palpe du maxillipède. Ce n’est d’ailleurs pas la première fois qu’une telle situation se présente comme l’ont en particulier déjà fait remarquer Hale (1925) et Brusca (1983).
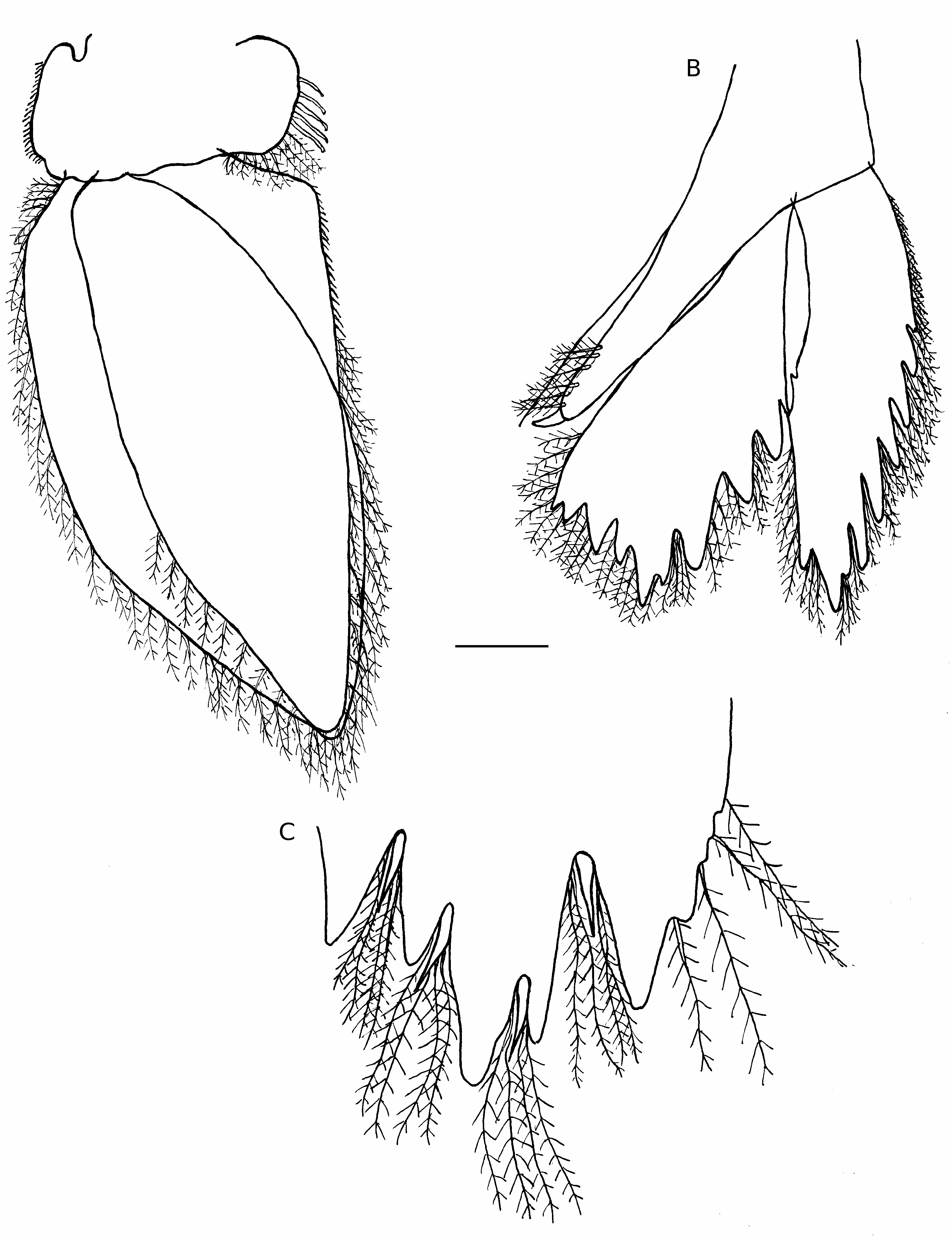
Aega angustata Whitelegge, 1901 View in CoL ( Figs 6 View FIG ; 7 View FIG )
Aega angustata Whitelegge, 1901: 203 View in CoL , 232-235, fig. 21a-f. — Hale 1925: 169-171, fig. 20a-f; 1940: 295. — Nierstrasz 1931: 182. — Barnard 1960: 94. — Moreira & Sadowsky 1978: 109. — Stephenson 1980: 153-155, figs 1-5. — Bruce 1996: 129, 134, 135.
Aega (Aega) angustata View in CoL – Brusca 1983: 10.
MATÉRIEL EXAMINÉ. — CHONDRICAL, Alis, pêche à la palangre, PAL 8, au large de l’Île des Pins, 22°54’71”S, 167°38’27”E, 325-348 m, au niveau de l’oeil d’un Squalus megalops (Macleay, 1881) View in CoL (955 mm; sélacien, Squalidae Bonaparte, 1834 View in CoL ), 31.I.2002, 1 JN082A, LT 26 mm, lt 7 mm ( MNHN-IS 5820 ). — PAL 1, Baie du Santal, Île de Lifou, 167°01’60”E, 20°53’30”S, 298-397 m, sur le ventre d’un Hexanchus nakamurai Teng, 1962 View in CoL (1664 mm; sélacien, Hexanchidae Gray, 1851 View in CoL ), 26.I.2002, 1 juvénile avec P7 encore rudimentaires JN001B, LT 9,5 mm, lt 3,0 mm ( MNHN-IS 5821 ). — PAL 10, au sud de la Nouvelle-Calédonie, 166°55’83”E, 23°01’06”S, 467-489 m, 1.II.2002, sur le corps d’une femelle de Squalus melanurus Fourmanoir & Rivaton, 1979 View in CoL (663 mm; sélacien, Squalidae View in CoL ), 1 juvénile (manca) avec uniquement les six premières paires de péréiopodes JN105A, LT 10,5 mm, lt 3,5 mm ( MNHN-IS 5822 ) GoogleMaps .
NOTES SYSTÉMATIQUES
Le mâle de cette espèce a été relativement bien décrit et figuré par Whitelegge (1901). Par la suite, la femelle a été décrite par Hale (1925) alors que plus récemment, Stephenson (1980), grâce à l’examen de deux spécimens récoltés en Nouvelle-Zélande, apportait quelques précisions complémentaires sur la morphologie du mâle, en particulier en ce qui concerne les pièces buccales. Toutefois, d’après Bruce (1996), les exemplaires de Stephenson appartiennent peut-être à l’espèce Aega komai Bruce, 1996 , dont l’auteur souligne également la ressemblance avec les espèces A. angustata et A. dofleini Thielemann, 1910 . Mais, pour pouvoir en décider sans ambiguïté, il serait indispensable d’examiner avec précision le pléotelson de ces deux spécimens, ce qui n’est pas possible sur les deux photographies produites par l’auteur.
Cette espèce, dont les antennes sont du type Aega (Aega) de Brusca (1983) , est surtout caractérisée par la présence d’un pléotelson très denticulé (comme d’ailleurs les uropodes), acuminé distalement et pourvu d’une abondante ornementation de soies plumeuses et de soies épineuses et d’un prolongement très développé au niveau du propodite des péréiopodes de la troisième paire et qui, associé au dactylopodite, simule presque une pince, ce qu’avaient déjà souligné Hale (1925) et Stephenson (1980). Pour Hale (1925), ce prolongement est d’ailleurs plus développé que chez aucune autre espèce du genre; mais l’auteur ne connaissait évidemment pas certaines autres espèces et en particulier l’espèce Aega chelipous décrite par Barnard (1960), à partir d’un spécimen femelle récolté à Madagascar sur Carcharinus sp. ; en effet, chez cette espèce, ce prolongement, terminé en une extrémité aiguë et non pas obtuse comme par exemple chez A. angustata ou A. komai , simule nettement plus une pince avec le dactyle du péréiopode 3. Parmi les espèces actuellement décrites, rappelons qu’un tel prolongement est présent uniquement sur les péréiopodes 3 chez A. angustata , A. chelipous , A. dofleini et A. komai .
Pour Hale (1940), comme pour Bruce (1996), A. angustata est voisine de l’espèce A. dofleini . Hale (1940) remarque toutefois que chez A. dofleini la lame frontale est tronquée alors qu’elle est arrondie chez A. angustata et que les antennes A1 sont très différentes (y compris le nombre d’articles). On peut ajouter, si l’on se base sur les illustrations de Thielemann (1910), que les uropodes et le pléotelson paraissent bien moins denticulés chez A. dofleini que chez A. angustata . Sur nos spécimens, on remarque en moyenne six indentations très nettes de part et d’autre de la pointe distale du pléotelson; ce dernier porte en outre une rangée de soies plumeuses avec une à deux soies épineuses au niveau de chacune de ces indentations.
Par rapport à A. komai , on peut souligner que nos spécimens sont certes assez voisins mais qu’il existe, entre ces deux espèces, en particulier dans la morphologie du pléotelson (nettement acuminé chez nos exemplaires et nettement tronqué chez A. komai ) et des uropodes, une différence si évidente qu’elle ne peut a priori laisser subsister aucun doute quant à leur distinction.
Taille
Mâle: 15 à 21 mm; femelle: 24 à 26 mm; juvéniles: 9,5 à 10,5 mm.
Hale (1940) signale deux spécimens de 10 et 29 mm, mais sans en préciser le sexe.
DISTRIBUTION
L’aire de répartition de cette espèce paraît bien définie puisqu’elle correspond au Pacifique sudouest. Compte tenu de la synonymie admise
A jusqu’à présent, nous pouvons indiquer qu’elle a été successivement signalée dans les localités suivantes: 1) Australie, Nouvelle Galles du Sud, Wata Mooli, à une profondeur de 98 à 107 m ( Whitelegge 1901), Eden, Botany Bay ( Hale 1925); 2) Australie et Tasmanie, Victoria, à une profondeur de 146 m, Gabo Islands; 3) Oyster Bay, « Tasmania Coast », Flinders Island, Bass strait, Great Australian Bight, 60 à 80 miles à l’ouest de Eucla et à 146 à 200 m de profondeur ( Hale 1940); et 4) Nouvelle-Zélande ( Stephenson 1980).
Cette espèce n’est pas citée par Bruce (1983) dans son étude sur les Aegidae d’Australie ; par contre, à l’occasion de la description de A. komai du Japon et afin d’établir une comparaison entre cette espèce et A. angustata, Bruce (1996) précise avoir examiné deux spécimens femelle de « Lakes Entrance, Victoria, Australia » et une femelle, non ovigère, LT 22 mm, en provenance de l’est de Sydney, Nouvelle Galles du Sud.
Nos spécimens étendent donc la distribution de cette espèce à la Nouvelle-Calédonie.
REMARQUES
Jusqu’à présent, cette espèce a été signalée: 1) sur un « Saw-fish » ( Hale 1925), probablement un sélacien Pristidae Bonaparte, 1838 , Pristis sp. d’après Moreira & Sadowsky (1978); 2) sur un requin dont l’espèce n’est pas précisée (« from a shark »; Hale [1940]); 3) sur un seul spécimen de « Grey-Spiny dogfish », Squalus blainvillei (Risso, 1827) , sur la peau, en arrière des nageoires pectorales, sans dommages tissulaires visibles ( Stephenson 1980).
Les spécimens de Nouvelle-Calédonie étendent et précisent la localisation possible de cette espèce sur trois autres sélaciens: Hexanchus nakamurai , Squalus megalops et Squalus melanurus .
Aega cf. deshaysiana ( Milne Edwards, 1840) ( Fig. 8 View FIG )
Rocinela deshaysiana Milne Edwards, 1840: 243 View in CoL . — Heller 1866: 22.
Rocinela deshayesiana – Studer 1883: 22. Aega deshayesiana – Schioedte & Meinert 1879: 360, 361, pl. VIII (Cym. II), figs 7-9. — Norman 1904: 434, pl. XII, figs 1-4, pl. XIII, figs 10, 11. — Richardson 1904b: 674; 1906: 821. — Thielemann 1910: 26, 27. — Nierstrasz 1918: 107, 108. — Bruce 1983: 782. — Rokicki 1984: 1-220, figs 17, 18; 1985: 95-122.
Aega deshaysiana – Richardson 1904a: 29. — Nierstrasz 1931: 182. — Hale 1940: 295; 1952: 27. — Moreira & Sadowsky 1978: 122. — Sivertsen & Holthuis 1980: 33, 34.
Aega (Aega) deshaysiana – Brusca 1983: 13-16, figs 1d-f, 2, 3. — Kensley & Schotte 1989: 117, fig. 51A.
Aega antillensis Schioedte & Meinert, 1879: 361 View in CoL , 362, pl. VIII (Cym. II), figs 10-13. — Richardson 1901: 520, 521; 1905: 167, 170, 171. — Thielemann 1910: 26, 27, figs 1, 2. — Nierstrasz 1918: 107, 108. — Hale 1925: 176-178, fig. 24a-h; 1929: 254, fig. 248; 1940: 295. — Barnard 1925: 389, 390. — Menzies & Frankenberg 1966: 5. — Schultz 1969: 189, 190, fig. 296a-d. — Kensley 1975: 39; 1976: 265, 290; 1978: 56-58, fig. 24A, B. — Moreira & Sadowsky 1978: 99, 108. — Brusca 1983: 13-16. — Kensley & Schotte 1989: 117.
Aega schioedteana Bovallius, 1885: 5-8 View in CoL , pl. I, figs 1-10. — Stebbing 1893: 349. — Brusca 1983: 13, 15.
Aega excisa Richardson, 1910: 11 View in CoL , 12, fig. 11. — Hale 1925: 178. — Brusca 1983: 13, 15.
MATÉRIEL EXAMINÉ. — CHONDRICAL, Alis, pêche à la ligne à beryx PAL 6, 167°04’04”E, 20°38’03”S, profondeur 600-681 m, dans l’évent d’un Squalus megalops View in CoL (sélacien, Squalidae View in CoL ), 28.I.2002, 1 JN019A, LT 29 mm, lt 10 mm (MNHN-IS 5823). — Pêche à la palangre PAL 7, au large de l’Île des Pins, 167°36’75”E, 22°59’80”S, profondeur 662-753 m, dans l’évent d’un Squalus cf. megalops View in CoL (850-900 mm), 31.I.2002, 1 juvénile (les P7 ne sont pas encore complètement développés) JN058A, LT 13 mm, lt 5 mm (MNHN-IS 5824). — Pêche à la palangre PAL 8, au large de l’Île des Pins, 167°38’27”E, 22°54’71”S, profondeur 325-348 m, dans l’évent d’un Squalus megalops View in CoL (955 mm), 31.I.2002, 1 juvénile JN082A, LT 10 mm, lt 3,5 mm (MNHN-IS 5825). — Pêche à la palangre PAL 11, au sud de la Nouvelle- Calédonie, 166°31’29”E, 22°37’50”S, profondeur 528-556 m, dans l’évent d’un Squalus megalops View in CoL (916 mm), 2.II.2002, 1 juvénile JN120A, LT 14 mm, lt 5 mm (MNHN-IS 5826).
Le statut de cette espèce est relativement complexe comme l’ont en particulier déjà souligné Sivertsen & Holthuis (1980) et Brusca (1983). Nous pouvons également indiquer que Bruce (comm. pers.) termine actuellement une révision très utile du « complexe Aega deshaysiana ». Pour nos spécimens de Nouvelle-Calédonie, nous préférons donc parler de Aega cf. deshaysiana .
Pour l’instant, la dernière étude précise parue sur cette espèce est celle de Brusca (1983) et c’est donc celle qu’il convient d’adopter ici, en particulier en ce qui concerne la synonymie. À cette occasion, il nous a cependant paru nécessaire de préciser certaines références bibliographiques.
Rappelons que c’est Nierstrasz (1918) qui a proposé de mettre en synonymie les deux espèces Aega deshaysiana et A. antillensis Schioedte & Meinert, 1879 , tandis qu’un peu plus tard, Hale (1925) a admis l’identité avec A. excisa . C’est par contre Norman (1904) qui a ajouté A. schioedteana Bovallius, 1885 , dans la synonymie de cette espèce; quant à sa ressemblance avec A. magnifica (Dana, 1853) , elle a été discutée par Brusca (1983).
NOTES SYSTÉMATIQUES
Cette espèce, dont les antennes sont également du type Aega (Aega) de Brusca (1983) , a été décrite très succinctement par Milne Edwards en 1840; elle a été plus récemment redécrite en détail par Brusca (1983), à partir de l’examen des types de Aega deshaysiana , A. antillensis et A. excisa , plus cinq spécimens supplémentaires, l’ensemble correspondant à une vaste aire géographique: Hawaï, Japon, Antilles, golfe du Mexique, Philippines, Méditerranée et Pacifique Est Tropical.
À l’heure actuelle, Aega deshaysiana est surtout distinguée par la nette échancrure que montrent les endopodites des uropodes sur leur bordure externe. Mais, comme l’avaient en particulier déjà précisé Richardson (1904b, 1906), Nierstrasz (1918, 1931), Hale (1925), Richardson in Hale (1925), Barnard (1925), Sivertsen & Holthuis (1980) et Brusca (1983), l’ornementation des divers appendices apparaît relativement variable d’une localité à une autre, au sein de ce que Brusca définissait déjà comme le « complexe » A. deshaysiana , A. antillensis et A. excisa . C’est en particulier le cas pour les antennules, les antennes, les maxillipèdes et les uropodes. Remarquons toutefois que de telles différences entre les spécimens d’ A. antillensis s.s., d’Afrique du Sud et de Saint Paul, permettent à Kensley (1976) de penser qu’il ne s’agit finalement peut-être pas de la même espèce.
Il ne fait donc aucun doute qu’il était nécessaire de revoir en détail cette question. C’est, en principe, ce que l’étude de Bruce (en préparation) s’attache à réaliser.
Taille
A. deshaysiana : juvénile 18-33 mm et mâle 24 mm ( Schioedte & Meinert 1879); femelle 47 et 52 mm ( Nierstrasz 1918), 33 mm ( Sivertsen & Holthuis 1980) et 18 mm ( Kensley & Schotte 1989); de sexe non précisé « environ 1 pouce » soit 27,5 mm ( Milne Edwards 1840), 57 mm ( Hale 1940) et 48 mm ( Hale 1952).
A. antillensis View in CoL : juvénile 36 mm ( Schioedte & Meinert 1879), 10 mm ( Hale 1925); mâle 30,5 et 37,0 mm ( Thielemann 1910), 48 mm ( Hale 1925, 1929); femelle 46 et 58 mm ( Thielemann 1910), 30 et 47 mm ( Hale 1925); de sexe non précisé 35 mm ( Richardson 1905; Schultz 1969), 40 mm ( Barnard 1925) et 45 mm ( Kensley 1978).
A. excisa View in CoL : femelle 36 mm ( Richardson 1910).
A. schioedteana : mâle 18,5 mm ( Bovallius 1885).
DISTRIBUTION
La localité type de cette espèce est la Méditerranée. Cependant, compte tenu de la synonymie admise jusqu’à présent, son aire de répartition peut paraître très étendue, mais rappelons à nouveau que plusieurs mentions sont certainement douteuses.
Il a été ainsi fait mention respectivement des localités suivantes:
– pour A. deshaysiana: Méditerranée ( Milne Edwards 1840) ; Adriatique ( Heller 1866); Palerme (« Panormum »), Sicile, Adriatique, à Fayal, Açores ( Schioedte & Meinert 1879); Cap Vert, 15°40’N, 23°5’8”W, à une profondeur de 70 m (« 38 fadem ») ( Studer 1883); Bonabai, Afrique du Nord, à environ 45 à 100 m (« 25- 55 fathoms ») de profondeur ( Norman 1904); Hawaï, Pailolo channel, entre Molokai et Maui Islands, Côtes nord ouest de l’Amérique du Nord ( Richardson 1904b) ; océan Indien, Seychelles ( Nierstrasz 1918); Japon, Hawaï, Australie, Seychelles, Natal, Afrique NW, Îles du Cap Vert, Açores, Méditerranée, Antilles, à une profondeur de 40 à 416 m ( Nierstrasz 1931) ; Nouvelles Galles du Sud , Byron Bay , Victoria , Gabo Island , Tasmanie , South Australia , Marsden point, Kangaroo Island, « Ontong », Java, près des Îles Salomon, près de la côte et à 146,5 et 366 m (« 80-200 fathoms ») ( Hale 1940) ; stn 113, 42°40’S, 148°27’30”E, Maria Island , Tasmanie, à 122 m de profondeur ( Hale 1952) GoogleMaps ; Île Tristan da Cunha, Atlantique Sud ( Sivertsen & Holthuis 1980); Hawaï, Pailolo channel, entre Molokai et Maui Islands (Expédition Albatross), Costa Rica, Eastern Pacific , Cocos Islands, Chatham bay, Golfe du Mexique, à une profondeur de 100 et 254 à 308 m ( Brusca 1983) ;
– pour A. antillensis : Cuba ( Schioedte & Meinert 1879); Cuba, Antilles, Cozumel, Havane, à des profondeurs de 298 à 422 m (« 163-231 fathoms ») ( Richardson 1901, 1905, 1906); Japon, Sagami Bay, Misaki, Île Hatsushima, Fukuura, à une profondeur de 92 m (« 50 fathoms ») ( Thielemann 1910); Antilles, Japon ( Nierstrasz 1918); Umvoti River, Natal, à une profondeur de 238 m (« 130 fathoms ») ( Barnard 1925); South Sea Islands, Nouvelles Galles du Sud, Tasmanie, South Australia, Fowler, S Bay, à une profondeur de 183 m (« 100 fathoms ») ( Hale 1925); Cuba, Antilles, Cozumel ( Menzies & Frankenberg 1966); Antilles, à 298-422 m de profondeur ( Schultz 1969); Afrique du Sud, à 200 m de profondeur ( Kensley 1975); Îles Saint Paul, sud de l’océan Indien ( Kensley 1976);
– pour A. excisa : Philippines, Jolo Light, N 82°E, 6.75 mi. (6°02’55”N, 120°53’00”E), à une profondeur de 340 m (« 186 fathoms”) ( Richardson 1910) GoogleMaps ;
– pour A. schioedteana: Adriatique ( Bovallius 1885) .
Sous réserve d’une révision de la synonymie cidessus, les spécimens que nous avons étudiés étendent donc l’aire de répartition du complexe Aega deshaysiana à la Nouvelle-Calédonie. On peut également souligner qu’ils ont été récoltés à des profondeurs de 600 à 750 m, c’est-à-dire aux profondeurs les plus importantes parmi celles qui ont été signalées jusqu’à présent.
REMARQUES
Comme pour la majorité des Aegidae actuellement connus, des détails éco-biologiques précis concernant A. deshaysiana sont rares.
Nous avons déjà signalé que les Aegidae ne sont pas de véritables parasites, mais des prédateurs qui ne se fixent que temporairement sur certains poissons ou sélaciens pour se nourrir. Aussi n’estil pas étonnant que des spécimens de cette espèce aient été parfois récoltés sur le fond, « in shells and corals » ( Richardson 1910), « on coarse sand » ( Kensley 1975) ou libres sur des bateaux de pêche ( Kensley 1976). Mais, à l’heure actuelle, cette espèce a également été signalée sur des sélaciens et peut-être sur des téléostéens: 1) sur « Basking Shark », Cetorhinus maximus (Gunnerus, 1765) ( Hale 1925) ; 2) sur un « Tiger Shark », au niveau du cloaque ( Hale 1940), alors que pour cette même référence, Moreira & Sadowsky (1978) indiquent peut-être par erreur « shark (species unknown) »; 3) dans l’estomac d’un « Cat Shark » ( Hale 1952), ce qui laisserait peutêtre penser soit que les Aegidae peuvent éventuellement être consommés par certains sélaciens soit qu’il s’agissait d’un spécimen qui était fixé sur un hôte avalé par le requin en question. Quant à Sivertsen & Holthuis (1980), ils signalent en avoir récolté un spécimen sur un bateau de pêche, mais non attaché, bateau avec lequel avaient été pris des « Snoek », Thyrsites atun (Euphrasen, 1991) et des « Five finger », Nemadactylus monodactylus (Carmichael, 1819) ; les auteurs pensent qu’il s’agit probablement d’un isopode parasite d’ Acantholatris , mais en faisant toutefois remarquer que Kensley (1976), dans son travail sur les Îles Saint Paul et Amsterdam, dit avoir examiné un spécimen d’ A. « antillensis » sur un Thyrsites atun , au niveau de l’Île Saint Paul.
En Nouvelle-Calédonie, il ne fait aucun doute que Aega cf. deshaysiana peut être couramment récoltée sur des sélaciens, en particulier sur Squalus megalops , ce qui rapproche nos observations de celles de Hale (1925, 1940), dans des localités légèrement différentes.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Aega Leach, 1815
| Trilles, Jean-Paul & Justine, Jean-Lou 2004 |
Aega (Aega) angustata
| BRUSCA R. C. 1983: 10 |
Aega (Aega) deshaysiana
| KENSLEY B. & SCHOTTE M. 1989: 117 |
| BRUSCA R. C. 1983: 13 |
Aega excisa
| BRUSCA R. C. 1983: 13 |
| HALE H. M. 1925: 178 |
| RICHARDSON H. 1910: 11 |
Aega deshaysiana
| SIVERTSEN E. & HOLTHUIS L. B. 1980: 33 |
| MOREIRA P. S. & SADOWSKY V. 1978: 122 |
| HALE H. M. 1952: 27 |
| HALE H. M. 1940: 295 |
| NIERSTRASZ H. F. 1931: 182 |
| RICHARDSON H. 1904: 29 |
Aega angustata
| BRUCE N. L. 1996: 129 |
| STEPHENSON A. B. 1980: 153 |
| MOREIRA P. S. & SADOWSKY V. 1978: 109 |
| BARNARD K. H. 1960: 94 |
| NIERSTRASZ H. F. 1931: 182 |
| HALE H. M. 1925: 169 |
| WHITELEGGE T. 1901: 203 |
Aega schioedteana Bovallius, 1885: 5-8
| BRUSCA R. C. 1983: 13 |
| STEBBING T. R. R. 1893: 349 |
| BOVALLIUS C. 1885: 8 |
Rocinela deshayesiana
| ROKICKI J. 1984: 1 |
| BRUCE N. L. 1983: 782 |
| NIERSTRASZ H. F. 1918: 107 |
| THIELEMANN M. 1910: 26 |
| RICHARDSON H. 1906: 821 |
| NORMAN A. M. 1904: 434 |
| RICHARDSON H. 1904: 674 |
| STUDER T. 1883: 22 |
| SCHIOEDTE J. C. & MEINERT F. 1879: 360 |
Aega antillensis
| KENSLEY B. & SCHOTTE M. 1989: 117 |
| BRUSCA R. C. 1983: 13 |
| KENSLEY B. 1978: 56 |
| MOREIRA P. S. & SADOWSKY V. 1978: 99 |
| KENSLEY B. 1976: 265 |
| KENSLEY B. 1975: 39 |
| SCHULTZ G. A. 1969: 189 |
| MENZIES R. J. & FRANKENBERG D. 1966: 5 |
| HALE H. M. 1925: 176 |
| BARNARD K. H. 1925: 389 |
| NIERSTRASZ H. F. 1918: 107 |
| THIELEMANN M. 1910: 26 |
| RICHARDSON H. 1901: 520 |
| SCHIOEDTE J. C. & MEINERT F. 1879: 361 |
Rocinela deshaysiana
| HELLER C. 1866: 22 |
| MILNE EDWARDS H. 1840: 243 |