
Adalatherium hui Krause,Hoffmann, Wible,and Rougier,2020
|
publication ID |
https://doi.org/ 10.1038/s41586-020-2234-8 |
|
persistent identifier |
https://treatment.plazi.org/id/C62D8780-DE3E-FFCB-7E53-2758FDCEFAE0 |
|
treatment provided by |
Plazi |
|
scientific name |
Adalatherium hui Krause,Hoffmann, Wible,and Rougier,2020 |
| status |
gen. et sp.nov. |
Adalatheriidae Krause,Hoffmann,Wible,and Rougier,2020,fam.nov. Adalatherium hui Krause,Hoffmann, Wible,and Rougier,2020 , gen. et sp.nov.
Etymology. Adàla (Malagasy),‘crazy’; therium (Latinized form of the Greek θηριoν), ‘beast’;the species name hui is in reference to the late Yaoming Hu for his contributions to our knowledge of early mammals. Holotype. UA 9030 ,skull and postcranial skeleton.
Type locality and horizon. MAD99-15 , Berivotra study area (northwestern Madagascar). Upper Cretaceous series (Maastrichtian stage, 72.1–66million years (Myr) ago),Anembalemba Member,Maevarano Formation,MahajangaBasin 18 .Additionalinformationonthegeological context is provided in the Supplementary Information.
Diagnosis. A. hui differs from all other known Mesozoic mammaliaforms in possessing quadrangular upper postcanine tooth crowns with four major cusps and three connecting perimetric ridges mesially,
lingually and distally that border—on threesides—a central valley that opens buccally; and lower postcanine tooth crowns with four major cusps arranged in a diamond pattern and connected by four perimetric crests,and a prominent mesiobuccal basin on the two distalmost lower
postcanines. The full diagnosis is provided in the Supplementary Information.
Cranium
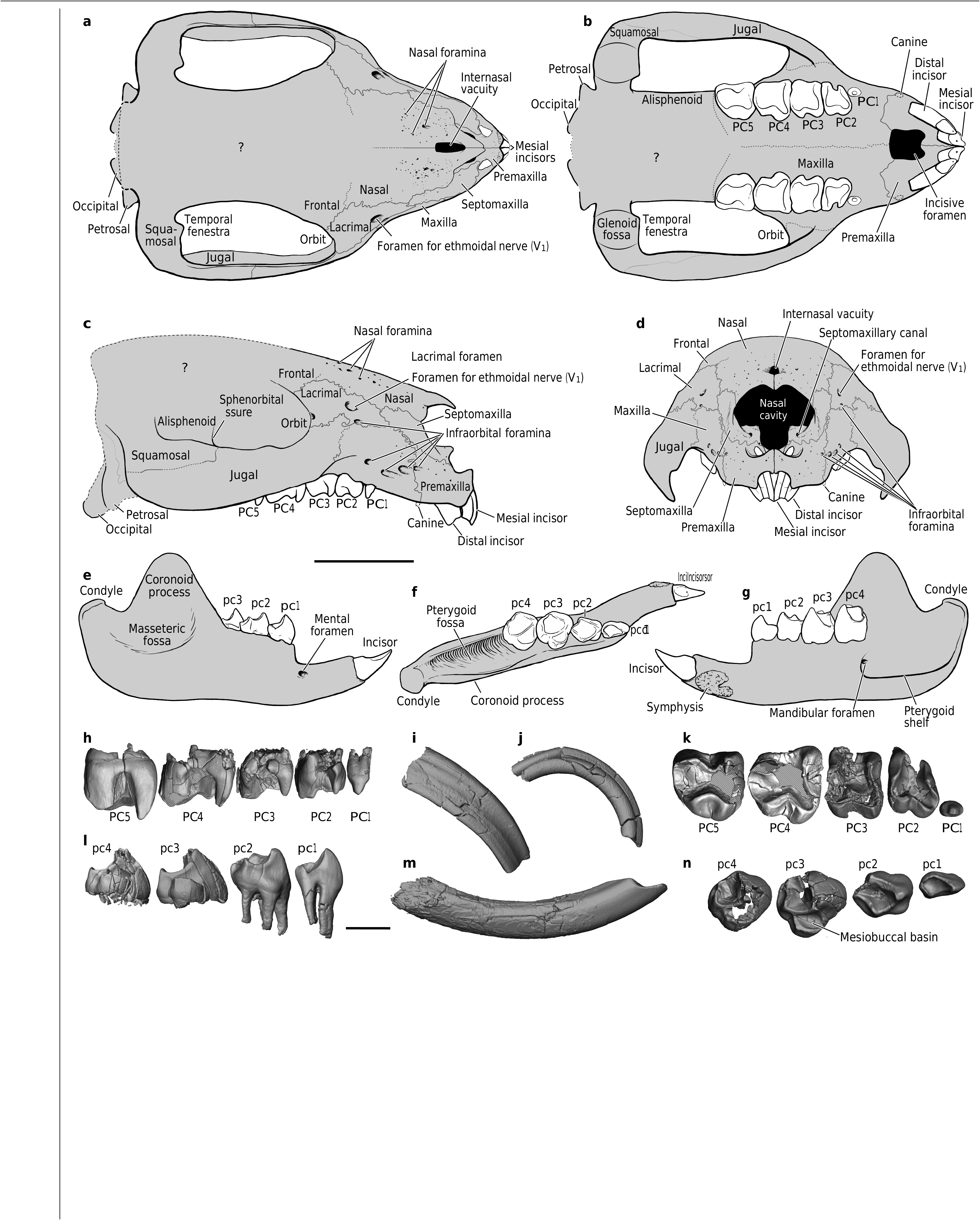
The cranium of Adalatherium reveals a mosaic of plesiomorphic and derived features ( Fig. 2 View Fig a–d, Extended Data Fig.3 View Fig ,Supplementary Videos 1–3). The presence of a very large internasal vacuity,five infraorbi-
tal foramina, a large foramen in the lacrimal that is not related to the nasolacrimal duct (probably for the ethmoidal branch of ophthalmic nerve (V 1) and associated vessels), numerous nasal foramina and a paranasal sinus that arises from the anterior vestibule of the nasal cavity are particularly unusual for mammaliaforms.The snout region shares several features with that of Vintana 3, 4, including the presence of a massive lacrimal bone that excludes the frontal from contacting the maxilla and a large septomaxilla with prominent posterodorsal and intranarial processes.By contrast, Adalatherium does not possess several autapomorphic features that are seen in Vintana , including a massive jugal flange and contact between the premaxillae and palatines. Although much of the posterior part of the cranium was severely damaged post mortem, the left inner ear of UA 9030 is partially pre- served and exhibits several features that were previously unknown among mammaliaforms (Extended Data Fig. 4 View Fig ). Most notably, the primary bony lamina is structurally different from that of therians in being single-layered instead of double-layered,and the branches of the cochlear nerve appear to have passed along the surface of—rather than within—the primary lamina.This unique morphology suggests that the primary bony lamina of Adalatherium evolved convergently with those of therian mammals.The cochleae of Adalatherium and Vintana are unique among mammaliaforms in possessing a secondary bony canal that parallels the cochlear ganglion canal,and probably enclosed a vascular network. Overall,the cochlear canal is curved through at least 210° and possesses, in addition to the primary bony lamina, the base of a secondary bony lamina,a cribriform plate,a well-developed cochlear ganglion canal and a separate canal for the lagenar nerve;this last feature is not present in Vintana 4.
Lower jaw
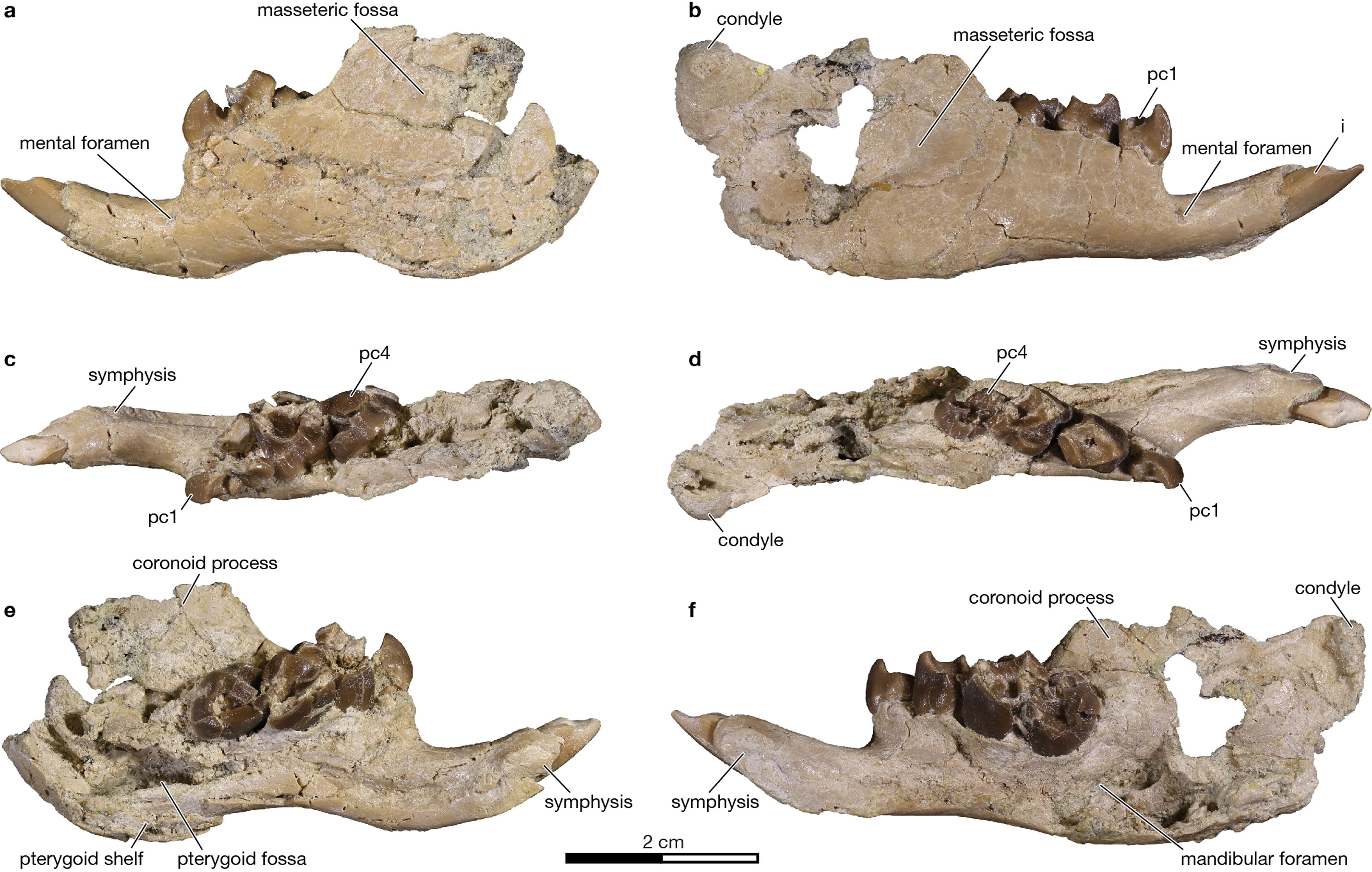
The lower jaw of Adalatherium is more complete than in any other known gondwanatherian,and is the first to preserve the ascending ramus of the dentary ( Fig.2 View Fig e–g, Extended Data Fig.5 View Extended ,Supplementary Videos 4–7).Among gondwanatherians,the horizontal ramus of Adalatherium is essentially identical to that of Sudamerica 19 but differs from that of Galulatherium 5, primarily in having a stepped differential in height between the diastema and the postcanine alveolar portion. The dentary of Adalatherium is short and deep,and bears a large diastema between the incisor and postcanine teeth,a prominent pterygoid fossa and shelf, and a masseteric fossa that extends anteriorly onto the horizontal ramus.There is no evidence of a postdentary trough, Meckelian sulcus,coronoid bone or angular process.In these features, the dentary of Adalatherium is similar to those found in members of Multituberculata , which is a largely Laurasian group. The dentaries of euharamiyidans differ from that of Adalatherium in possessing an angular process,a coronoid bone and—according to ref. 20 —a ‘reduced’ postdentary trough (although this is disputed in ref. 21). The dentary of Haramiyavia is much more plesiomorphic than those of both Adalatherium and euharamiyidans in retaining a long and shallow horizontal ramus,a fully developed postdentary trough and Meckelian sulcus, and in lacking a pterygoid fossa and shelf 22. The masseteric fossa of Adalatherium is positioned relatively high dorsally on the dentary,an apparent autapomorphy.
Dentition
The dentition of Adalatherium is unlike that of any known Mesozoic mammaliaform ( Fig. 2 View Fig h–n, Supplementary Video 8 for upper postcanines). There are two very large,open-rooted upper incisors, each of which bears a buccally restricted band of enamel. The size, shape and positional relationships of the upper incisors are very similar to those discerned from the alveoli of Vintana 3, 4. The presence of upper canines in Adalatherium is indicated by tiny elliptical alveoli that are separated mesially from the incisors and distally from the postcanines by sizeable diastemata. The first upper postcanine is a small, simple, two-rooted premolariform tooth. The four more-distal postcanines are each supported by five or more roots and are unique among Mesozoic mammaliaforms in bearing four major cusps connected by ridges mesially, lingually and distally that border—on three sides—a central valley that opens buccally.
The single lower incisor is large,curved and open-rooted,and bears enamel that is largely restricted to the buccal surface.In these features, the lower incisor resembles those known for other gondwanatherians (except for the enamel-less condition in Galulatherium 5). Each of the four lower postcanines has four cusps connected by prominent crests, forming a diamond pattern.The most mesial cusp dominates the crown on all four teeth. The second lower postcanine bears a mesiobuccal bulge,which is developed into a prominent basin on the penultimate and ultimate lower postcanines.The first lower postcanine has two roots, whereas each of the more distal postcanines has at least four. The enamel microstructure of Aadalatherium consists of relatively plesiomorphic ‘normal’ radial enamel (Extended Data Fig. 6 View Extended ), typi- cal of several gondwanatherians from the Late Cretaceous epoch and Palaeogene subperiod of Argentina 3, 4. It is unlike the ‘modified’ radial enamel,with pronounced inter-row sheets of interprismatic matrix, documented for other gondwanatherians( Lavanify and Vintana ) from the Late Cretaceous epoch of India and Madagascar 3, 4.
Postcranial skeleton
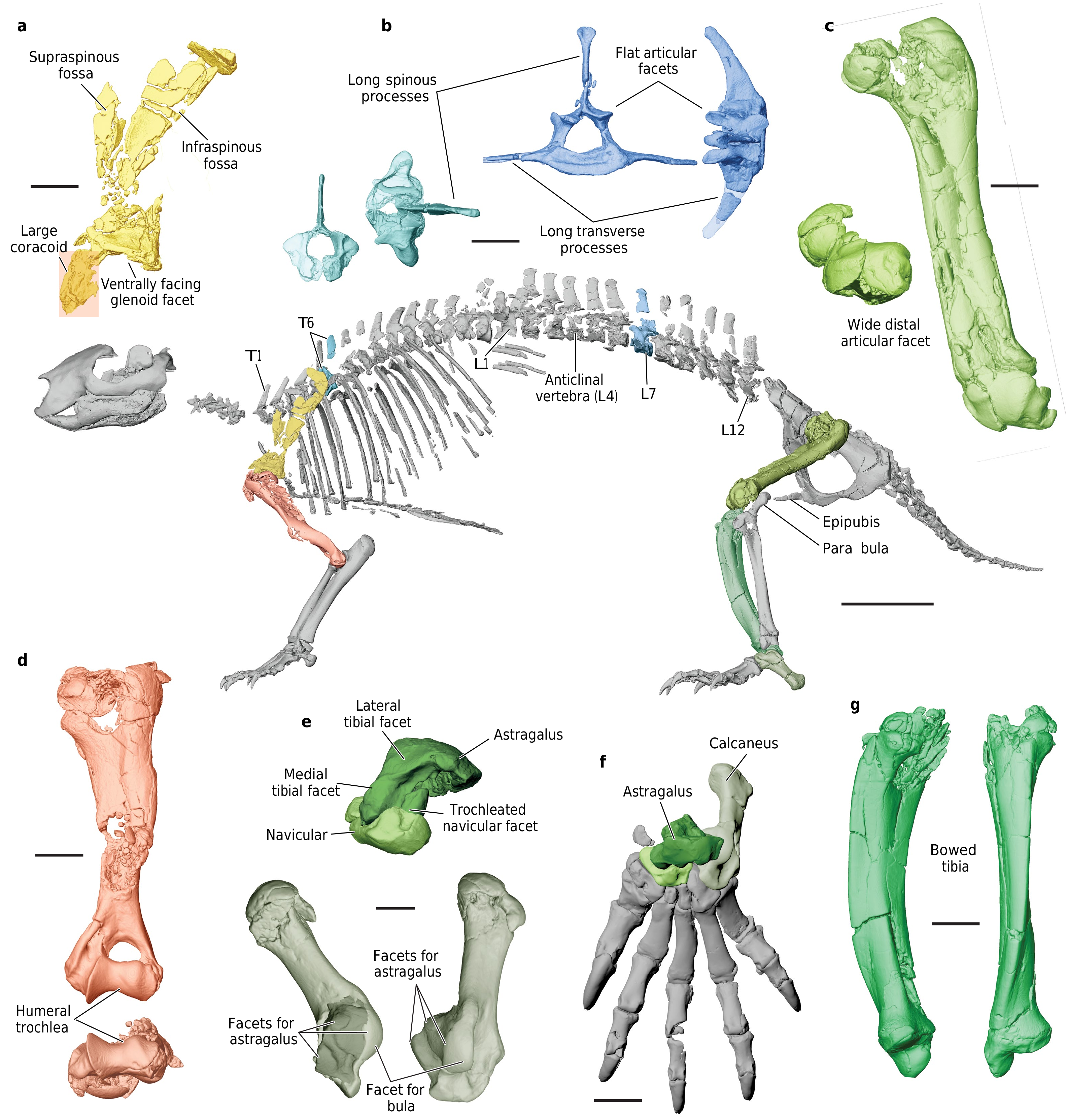
UA 9030 includes the only postcranial material assigned to a gondwanatherian,and Adalatherium isonlythefourthMesozoicmammaliaform from Gondwana represented by articulated postcranial remains 1. The postcranial skeleton exhibits a number of unusual features,including an anteroposteriorly bowed and mediolaterally compressed tibia,a trochleatedsurfaceonthedistalastragalus,alargenumberoftrunkvertebrae (at least16thoracic and12lumbar vertebrae),and a short tail(24vertebrae, almost all wider than they are long) ( Figs.1 View Fig , 3 View Fig , Extended Data Figs.1 View Fig , 7 View Extended ,8). The long spinous and transverse processes of the thoracic and lumbar vertebraesuggestthepresenceofenhancedepaxial(back) musculature. In the pectoral girdle,a procoracoid bone is absent but a separate coracoid is well-developed ( Fig.3a View Fig , Extended Data Fig.9a,b View Extended ). The forelimbs had a moderately parasagittal posture,as indicated by the ventrally directed glenoid fossa and the well-developed humeral trochlea( Fig.3a,d View Fig ,Extended DataFig.8a,b).Bycontrast,theasymmetricalmedialandlateralcondyles of the femur are suggestive of a more sprawled hindlimb posture.Other notable features of the pelvis and hindlimbs include the presence of a largeobturatorforamen (similarinsizetothoseoftherians),anepipubic bone and a large,separate parafibula ( Fig.3 View Fig ,Extended Data Fig.9c, d View Extended ).
Phylogenetic relationships
Our phylogenetic analysis, performed using 84 cynodont taxa and 530 morphological characters, places Adalatherium within Gondwanatheria, which in turn is placed within Allotheria as the sister taxon to Multituberculata (Extended Data Fig. 10 View Extended ,Supplementary Information). Adalatheriidae (as solely represented by Adalatherium ) is recovered as more derived than Ferugliotheriidae and stemward relative to Sudamericidae .
Previous phylogenetic analyses that include the recently discovered Early Cretaceous purported haramiyidan Cifelliodon 23, 24 advanced the idea that gondwanatherians are nested within Eleutherodontida, basal to the purported Early Cretaceous hahnodontids Hahnodon 25 and Cifelliodon 23. Although Hahnodon was not included in our analysis because it is represented by only one (or possibly two)isolated teeth 25, 26, Cifelliodon is recovered at the base of Allotheria, which also includes Euharamiyida and ‘ Multituberculata + Gondwanatheria’.Our analysis places the haramiyidans Haramiyavia and Thomasia —along with the poorly known taxon Megaconus —outside of Mammaliaformes,with no close relations to allotherians. This finding is in contrast to previous analyses for Vintana 3 and for Jurassic euharamiyidans 27 – 29.
Evolution in isolation
Amongmammals,themostobviousandquantifiableinfluencesofevolving on islands are those related to body size.This observation has led to the articulation of the ‘island rule’,which states that—evolutionar- ily—small mammals on islands increase,and large mammals decrease,in size 11, 30.Inaddition,evolutionininsularenvironmentsisthoughttoresult in changes in anatomy,physiology,behaviour and life-history strategies, and (at the faunal level)relatively low species richness,taxonomic imbalance,high endemism and a general level of ‘primitiveness’ 6, 12. Although somewhat controversial and clearly not ubiquitous 31, the island rule is generally established as a pervasive pattern 9, 11, 32. Examples of insular ‘dwarfism’ and ‘gigantism’ from Pliocene, Pleistocene and Holocene epochs abound 10, 11, 15, including from Madagascar (pygmy hippopotamuses 33 andgiantlemurs 34). ExamplesfromearlierintheCenozoiceraare relatively sparse 14, 16, 17 and the effects of long-term isolation are extremely poorly documented for Mesozoic mammaliaforms.
Among Mesozoic mammaliaforms,adaptations related to evolution in isolation have been most notably claimed for two island environments,both from the latest part of the Cretaceous period(Maastrichtian age): (1) the gondwanatherian Vintana from Madagascar 3 and (2) the multituberculates Barbatodon and Litovoi from the archaic ‘Hațeg Island’ 35, 36 (now part of Romania). Whether Barbatodon and Litovoi were part of a fauna that developed unique adaptations attributable to evolution in an insular environment is questionable—as is whether Hațeg Island was even an island (Supplementary Information).Because of its completeness and undoubted existence in an insular environment, the skeleton of Adalatherium provides an opportunity to examine evolution in isolation among Mesozoic mammaliaforms.
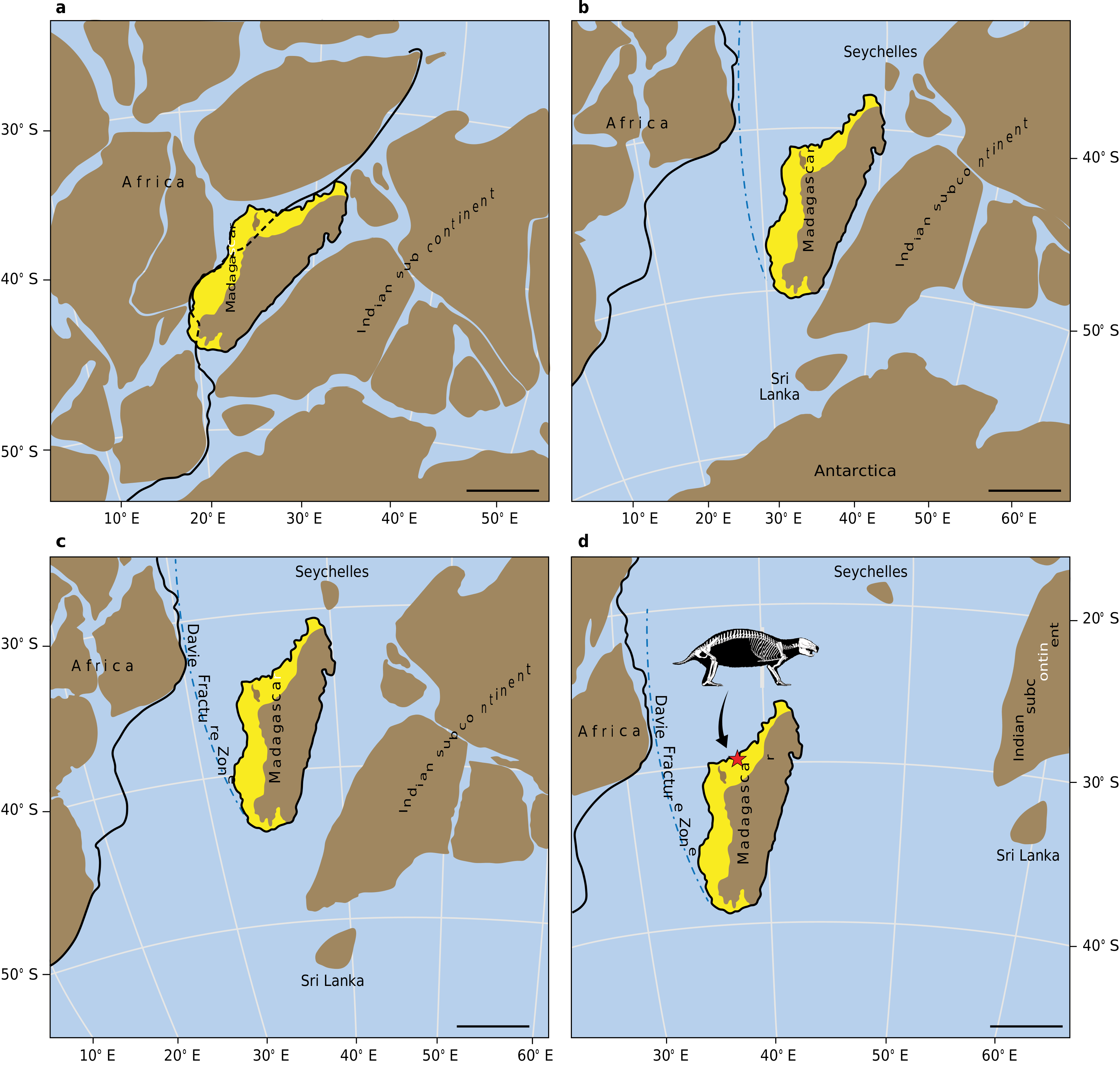
Madagascar separated from the Indian subcontinent and the Seychelles about 88 Myr ago 37, 38 ( Fig. 4 View Fig ). As a result,after separation, the obligate terrestrial taxa of Madagascar evolved in complete isolation and the only taxa that gained access to the island subsequently were flying, swimming or rafting forms that were able to disperse across considerable marine barriers 38 – 41. Madagascar remains today a large, isolated continental block that is topographically high, geotectonically stable and at a minimum distance of 430 km from the closest mainland (Africa).
The postcranial skeleton of UA9030indicates that Adalatherium (and perhaps other gondwanatherians) was neither volant nor aquatic;it was an obligate terrestrial form that was relatively less capable of dispersal across marine barriers and more likely to have evolved on Madagascar. There are at least two other gondwanatherians that lived on Madagascar contemporaneously with Adalatherium : Lavanify (based on two fragmentary isolated teeth 42 that are insufficient to be informative in the current context) and Vintana (based on a complete cranium 3, 4). The cranium and upper postcanine dentition of Vintana exhibit several features that are unknown among Mesozoic mammaliaforms,but the number of such features in Vintana are far fewer than those for Adalatherium (based on a complete skeleton).Furthermore,given its deeply nested phylogenetic position within Allotheria (Extended Data Fig. 10 View Extended ), several additional features of Adalatherium clearly evolved convergently with those of non-allotherian mammals—particularly therians (Supplementary Information). Considered together, and in concert with the palaeogeographical history of Madagascar,we hypothesize that the unusual morphological attributes of these two gondwanatherians are due to long-term evolution in isolation in an island environment, paralleling the cases adduced for various mammals on Mediterranean islands during the Neogene subperiod 6 – 8, 10. Furthermore,both Adalatherium and Vintana are large,being among the largest mammaliaforms known from the entire Mesozoic era of Gondwana. Vintana , which is even larger than Adalatherium , is the second-largest known Mesozoic mammaliaform globally and the largest from Gondwana 3, 4. Although the fossil record of Mesozoic mammaliaforms (especially from Gondwana) is too poor to conclusively establish that the large size of Vintana and Adalatherium reflects island gigantism,such a conclusion is consistent with the island rule 6 – 13, 30.
The vertebrate fauna associated with Adalatherium (Supplementary Table 2) also exhibits unique characteristics reflective of its relictual nature.Forinstance,relativelyhighnumbersofderivedfeaturesareseen in other terrestrial members of the latest Cretaceous fauna of Madagascar,including the ceratophryid frog Beelzebufo ,the crocodyliform Simosuchus and the theropods Majungasaurus and Masiakasaurus (ref. 38 and references therein).None of these genera is known from any other landmass,which again attests to high endemicity (although the very poor Mesozoic fossil record from Gondwana must be acknowledged in this regard).Furthermore,many of these forms had ghost lineages that extend back to or before theEarly–Late Cretaceous boundary (100 Myr ago) 43, thus suggesting that their ancestors had probably arrived on Indo-Madagascar before its separation from other Gondwanan landmasses(that is,via a deep-time vicariance event rather than overwater dispersal).Despite originating in different ways,we conclude that the latest Cretaceous insular vertebrate fauna of Madagascar was probably as unique relative to mainland faunas as it is today.
The currently known latest Cretaceous (Maastrichtian) vertebrate fauna of Madagascar, assembled as an island fauna over the course of more than 20 Myr (around 88–66 Myr ago),became completely or nearlycompletelyextinct—presumablytheresultoftheend-Cretaceous bolide impact and/or the penecontemporaneous nearby volcanic eruptions that resulted in the Deccan Traps of India 44. Thereafter,assembly of a Madagascan fauna began afresh.The establishment of a new vertebrate fauna required the arrival of transoceanic dispersers,primarily from Africa,that encountered habitats that were largely to completely devoid of Maastrichtian antecedents 38 – 41.

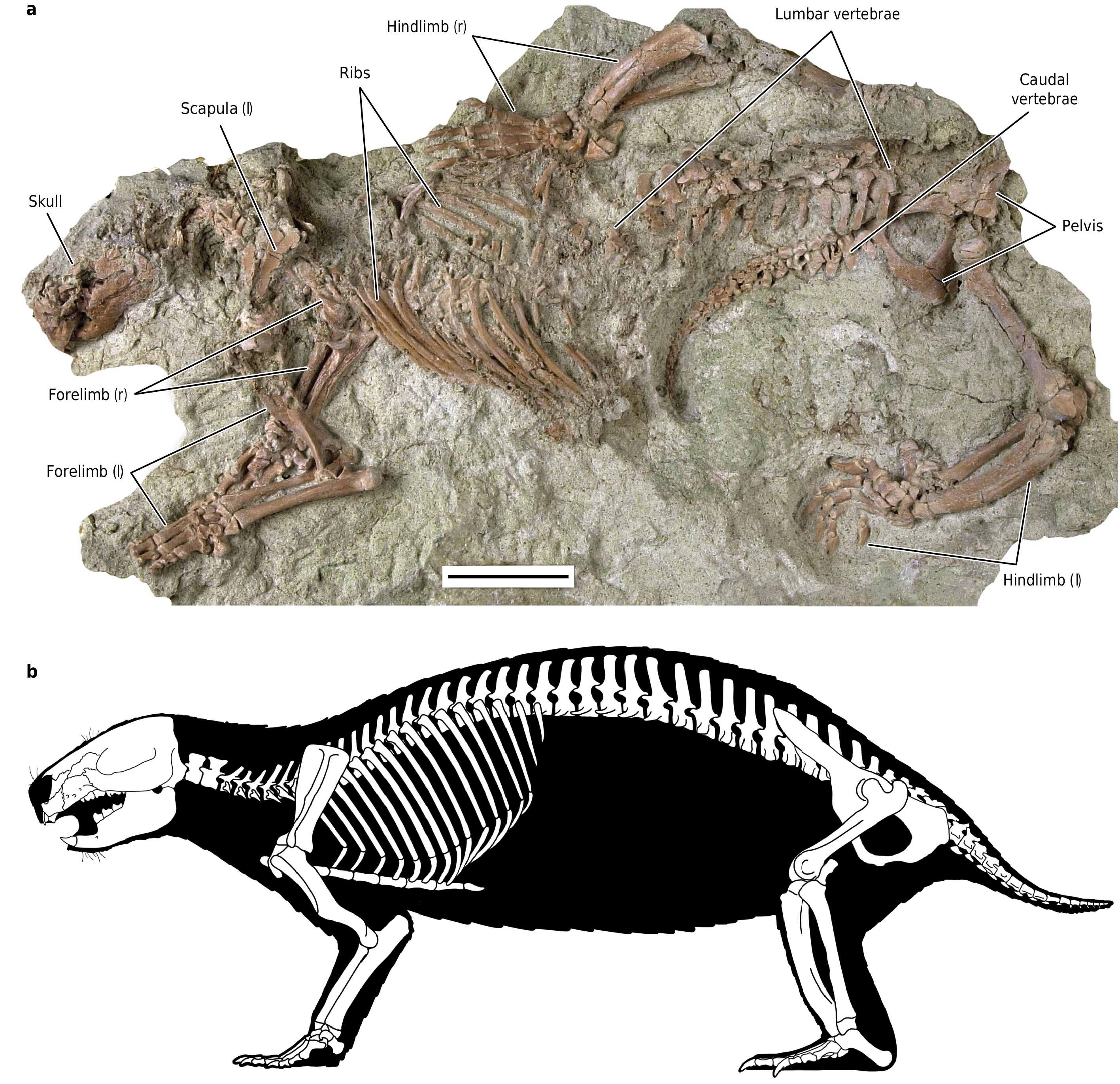
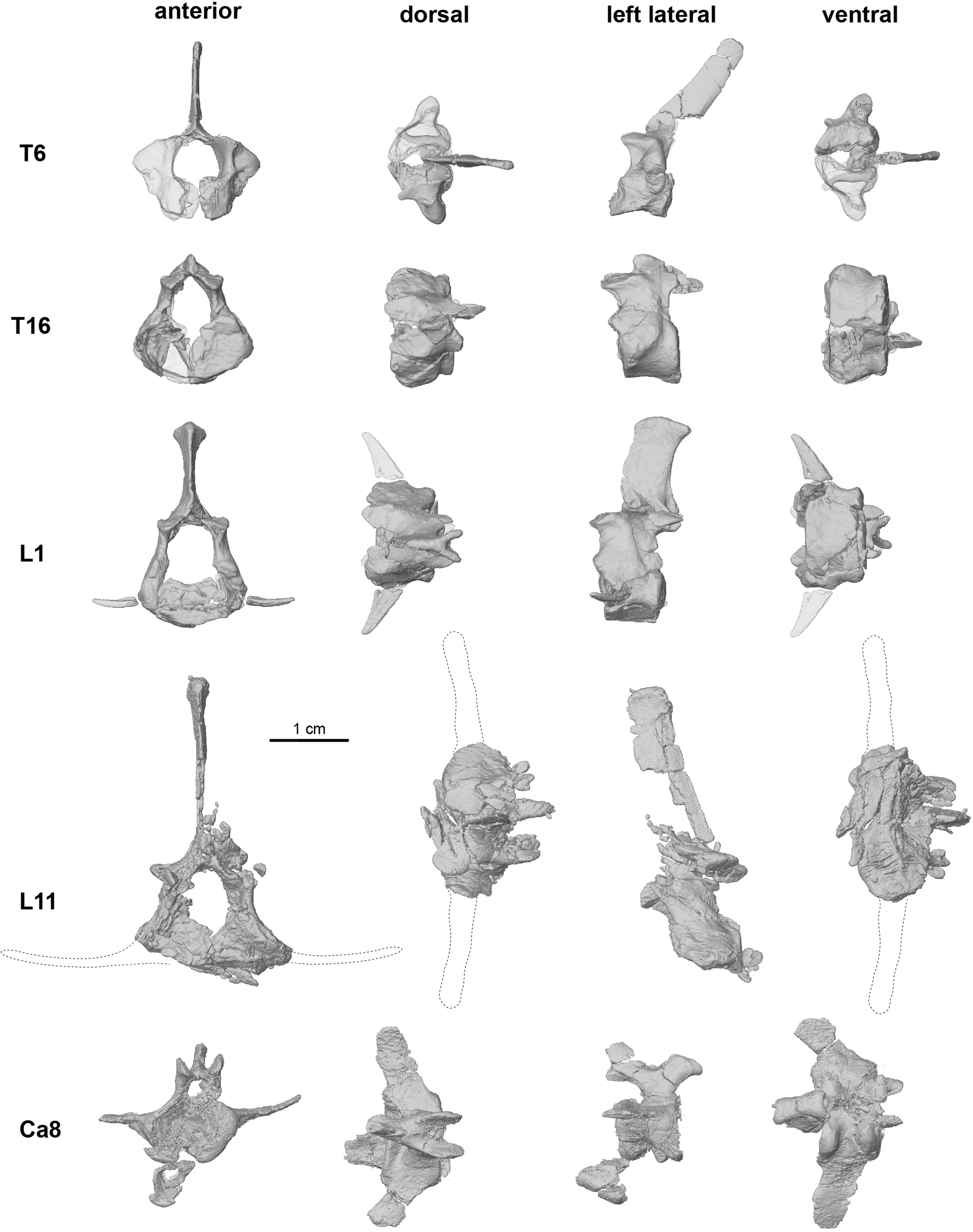
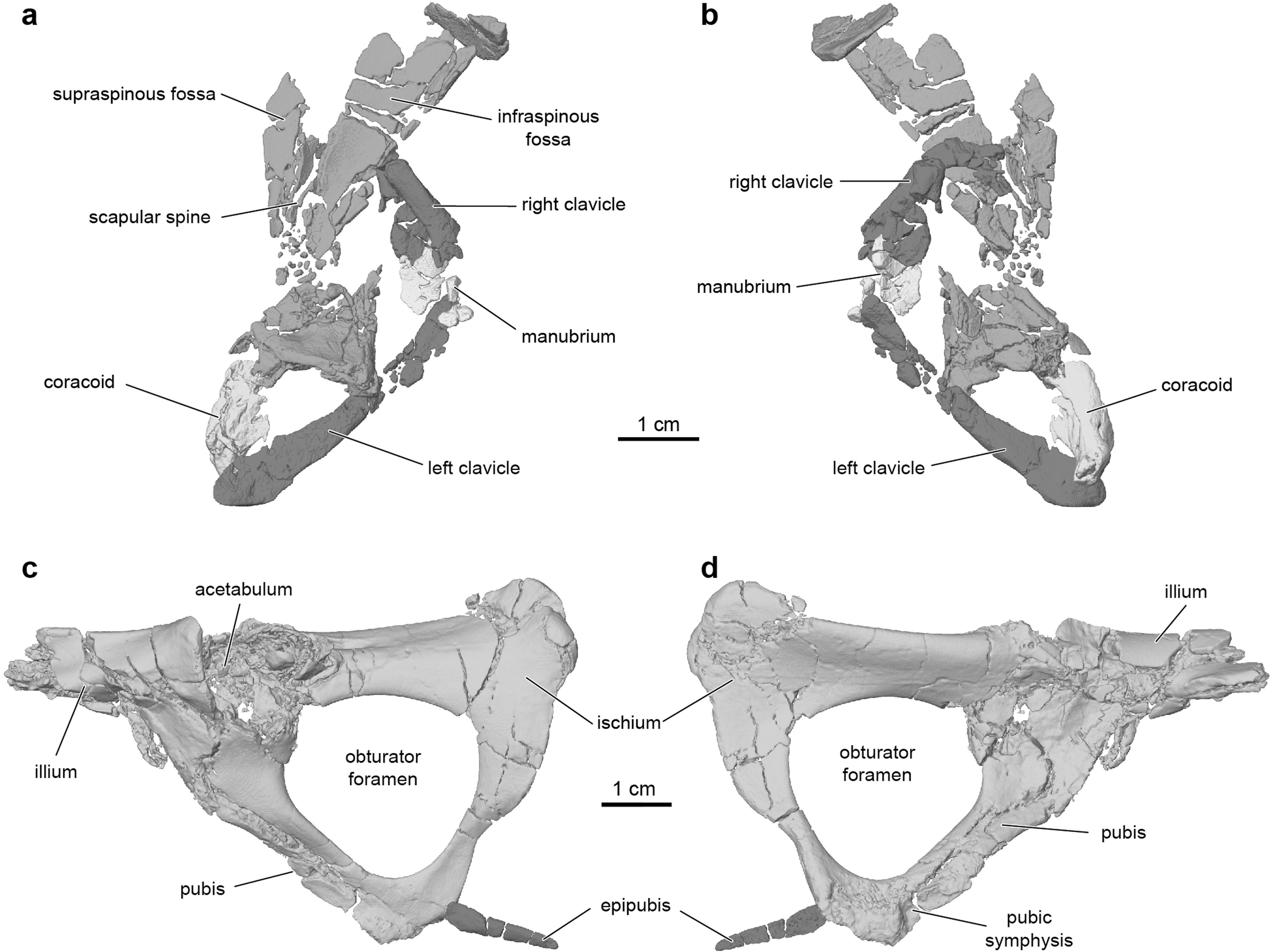
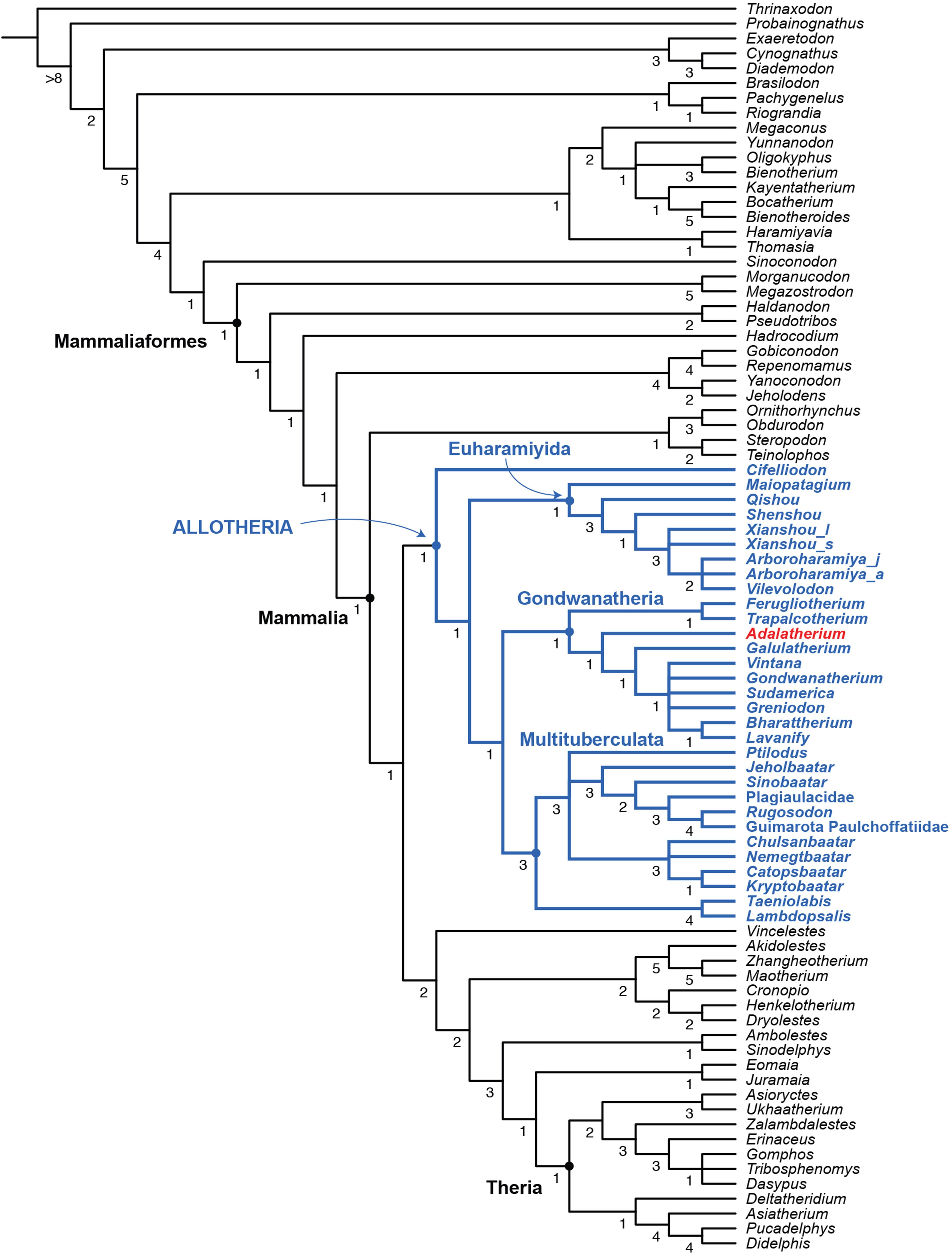
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Family |
|
|
Genus |