
Abyssocladia microstrongylata, Vacelet, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4767.2.3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:6A6A15AA-9B6D-4168-8857-29E183D965B9 |
|
DOI |
https://doi.org/10.5281/zenodo.3798812 |
|
persistent identifier |
https://treatment.plazi.org/id/0390B806-9829-FB5C-FF10-FBCDFAD1FEBA |
|
treatment provided by |
Carolina |
|
scientific name |
Abyssocladia microstrongylata |
| status |
sp. nov. |
Abyssocladia microstrongylata View in CoL sp. nov.
( Figs 5–6 View FIGURE 5 View FIGURE 6 )
Material examined. Holotype MNHN-IP-2015-1471 ( Fig 5A View FIGURE 5 ), KANACONO Expedition , New Caledonia (MEOW), beam trawl CP4754, 25/08/2016, 23°22’S 167°54’E, 1009–1019m. GoogleMaps
Paratype 1 MNHN-IP-2015-1472. Same data as holotype. A specimen broken in two fragments, with holdfast base.
Paratypes 2 MNHN-IP-2015-1473. Same data as holotype. A specimen in two fragments.
Paratypes 3 MNHN-IP-2015-1474 ( Fig 5B, C View FIGURE 5 ), several specimens or fragments, without the attachment base, two of them with a short papilla. KANACONO Expedition, New Caledonia, Station beam trawl CP4751, 25/08/2016, 23°21’S 167°57’E, 946–998 m,
Additional specimens. Several specimens or fragments, Station CP 4753 23°18’S 167°57’E 758–952m, 25/08/2016. Station CP 4754, 25/08/2016, 23°22’S. 167°54’E, 1009–1019 m (with holotype and paratypes 1 and 2).
Diagnosis. Cladorhizidae in a single axis, with bottle-brush filament arrangement and a few buds, three classes of styles, arcuate cleistochelae, arcuate isochelae, abyssochelae, sigma and sigmancistra, and a thick coating of microstrongyles on the axis and on the filaments.
Description. The sponge has a straight or slightly curved single axis, flattened in one plane, bearing numerous thin filaments perpendicularly arranged in two lateral rows ( Fig 5 View FIGURE 5 ). It is attached by a small, enlarged base. The most complete specimen attains 90 mm in length, but most are fragments devoid of attachment base or apical part. The axis is slightly flattened, 0.8–2 mm wide by 0.5 mm thick. In some specimens, the basal part is devoid of lateral filaments or has only very short filaments; this nude basal part attains 30 mm in one specimen. The filaments, 4 mm in maximum length, 150–250 µm in diameter at the base, with a spacing of 175–250 µm, are preserved only in places and are often reduced to the spicule axis. The attachment base, 4–5 mm in diameter, is very hard and includes a few rocky fragments.A few specimens have one or two short buds ( Figs 5B, C View FIGURE 5 ), 1–4 mm long with an axis 0.5 mm in diameter, ending in an enlarged button 1–2 mm in diameter. The consistency is rigid, but the sponge breaks easily. The color is brown in ethanol, with reproductive elements appearing darker in transparency, when present ( Fig 5C View FIGURE 5 ).
Skeleton. In most specimens the main skeleton is a thick axis in the stem, 750–1250 µm in diameter, composed mainly of long, fusiform styles or substrongyles 1 arranged in parallel, and also including a few styles 2. Some substrongyles 3, similar to the substrongyles 3 of the attachment base, are irregularly arranged around the main axis. These spicules are present in highly variable number in this position: they are sometimes absent, but may be very numerous in specimens or parts of specimens with stems attaining 2 mm in width in which the main axis of long styles is only 375–400 µm thick ( Fig 5D View FIGURE 5 ). The axis of the filaments, 120–200 µm in diameter, is made of the same styles 1, which are as long as those of the main axis, but generally less fusiform in shape. The filaments are implanted in the main axis by a basal part, in which the spicules are slightly diverging and cross the base of the opposite filament ( Figs 5D, E View FIGURE 5 ). The end of the filaments appears fragile and most are broken. In a few cases, however, the filament ends in a bunch-like bouquet of a few diverging styles, 300–400 x 5–6 µm. Both stem and filament axes are covered by a dense feltwork of microstrongyles, up to 70–100 µm thick on the stem, 20–30 µm thick along the filaments ( Fig 5E View FIGURE 5 ); this dense coating is preserved only in places. Arcuate cleistochelae 1 (55–70 µm), abyssochelae 2 (30 µm), sigmas and sigmancistras are present in the stem, dispersed without order, but mostly abundant between the stem axis and the feltwork of microstrongyles.
The enlarged attachment base, 4–5 mm in diameter and including a few rocky fragments, is made of a dense, hard feltwork of thick styles or substrongyles 3, some remarkably short and thick. Long styles are absent from the attachment base. The microscleres include sigmancistras, a few microstrongyles which are considerably less abundant than in the other parts of the sponge and abundant abyssochelae 2 similar to those of the other parts but generally smaller (17–22 instead of 25–32 µm).
The few buds observed have a cover of a feltwork of microstrongyles near the base, and a short axis of styles diverging near the middle in an enlarged button made of diverging fascicles, sometime with an irregular coating of arcuate cleistochelae 1 at the end, most of them with the alae and teeth outwardly directed. The microstrongyles are present at the end of the button, but do not form a dense feltwork.
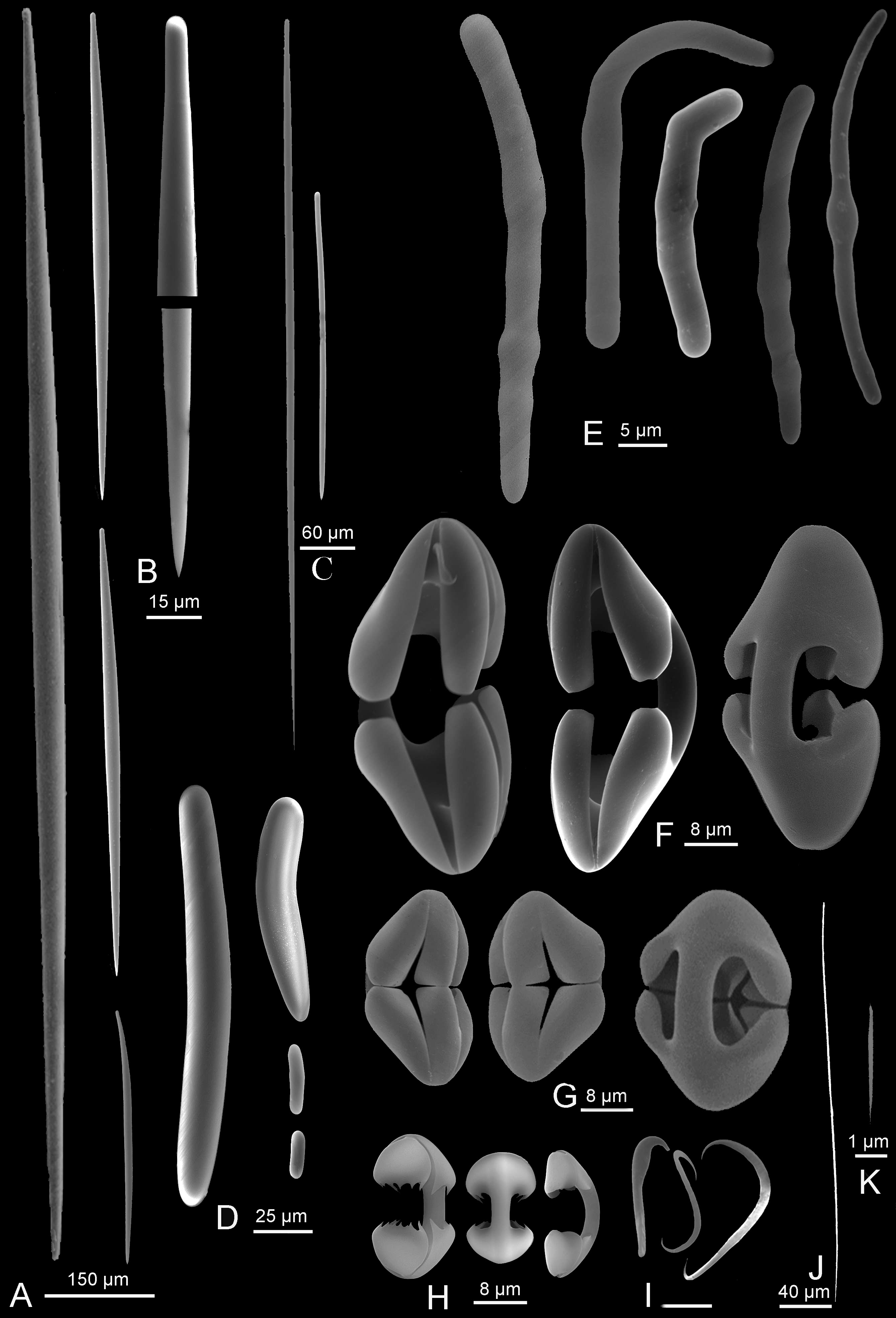
Spicules ( Fig 6 View FIGURE 6 ). There are many transitions between the megascleres, which are difficult to characterize.
Styles or substrongyles 1 ( Fig 6A, B View FIGURE 6 ) of the axis of the main stem and of the filaments: straight or slightly curved, highly variable in size, the longer ones strongly fusiform and often with a blunt tip, especially in the main axis: 338–1800 x 15–45 µm.
Styles 2, ( Fig 6C View FIGURE 6 ) mixed with styles 1 in the axis and in the filaments, straight and only weakly fusiform: 150–875 x 5–15 µm. Some, generally thinner and more uniform in size, diverge in bouquet at the end of the filaments: 300–400 x 5–6 µm
Styles or substrongyles 3 ( Fig 6D View FIGURE 6 ), thick and slightly curved, the tip being in general similar to the head. The longer ones are slightly fusiform and thus transition to styles 1: 18–580 x 6.4–35 µm. They are more numerous near the base, especially the smaller ones, which may be only 18 x 6.4 µm.
Thin styles ( Fig 6J View FIGURE 6 ), straight or slightly curved near the head, 220–290 x 2–2.5 µm. Found only in embryos.
Microstrongyles or microsubstrongyles ( Fig 6E View FIGURE 6 ), abundant everywhere except in the attachment base, are more or less curved, sometimes strongly angular, with a smooth surface or more often bearing a variable number of swellings. The two ends are often slightly unequal: 17–50 x 2.5–5 µm. Some spicules are intermediates with the styles or substrongyles 3, and although less numerous than the small microstrongyles with swellings are always present.
Arcuate cleistochelae ( Fig 6F View FIGURE 6 ): curved shaft, opposite central tooth and alae separated by a short distance: 55–70 x 20–35 µm, very rarely 50 or 80 µm in length.
Abyssochelae ( Fig 6G View FIGURE 6 ): strongly curved shaft, opposite central tooth and alae which are very thick and nearly touching or even in contact: 16–40 x 15–20 µm, with a height / width ratio varying from 1.2 to 1.6.
Arcuate isochelae ( Fig 6H View FIGURE 6 ): found only in a few specimens with embryos and probably juvenile stages, with large well separated teeth and most often bearing short, acute spines: 17–35 µm, 17.5–18 µm in holotype.
Sigmancistras ( Fig 6I View FIGURE 6 ), generally in two sizes, 7.5 µm and 12.5 µm (with the smaller being more numerous).
Sigmas ( Fig 6I View FIGURE 6 ), 12–20 µm, possibly in two sizes, the larger being very rare and not observed in some specimens.
Microraphides ( Fig 6K View FIGURE 6 ) found only in some embryos, very small: 4 x 0.08 µm.
Living tissue and reproductive elements. The living tissue is poorly preserved and is mainly observed in the reproductive elements. No living tissue is visible on the surfaces of the axis or the filaments. Two types of bodies which are most probably reproductive elements are present in the middle of the axis of several specimens, located in a cavity between the spicules ( Figs 5 View FIGURE 5 E–H).
The largest type, very likely embryos at diverse development stages, are roundish or ovoid with an enlarged pole, varying from 500 µm in diameter to 625 x 320 µm. Their cellular content may be composed of cells about 15 µm in diameter with numerous round inclusions of 2 µm in diameter, but these cells cannot be clearly distinguished given the quality of histological fixation and the thickness of the polished slides available. The tissue of these probable embryos contains numerous spicules, varying according to the maturation stage. Some have only isochelae 25–30 µm long with a very thin shaft. Others have isochelae of the same size, but more similar to the abyssochelae of adult tissue ( Fig 5G View FIGURE 5 ), and numerous sigmas 12–13 µm long. Others ( Fig 5H View FIGURE 5 ) have one pole devoid of spicules and the other part with abyssochelae 25–30 µm, sigmas and fascicles of thin styles 220–250 x 2–2.5 µm as shown in Figs G, J. Sigmancistras were not observed in these embryos. It is highly probable that arcuate isochelae with large spinose teeth ( Fig 6H View FIGURE 6 ) which have been observed in a few specimens only, are located in maturing embryos, but they were not observed in the few sections examined.
Smaller inclusions ( Figs 5F, G View FIGURE 5 ), 40–60 µm in diameter, likely sperm cysts in diverse stages of maturation, were observed also in the center of the stem of the same sponges, but in a different location than the embryos. They contain either poorly distinct cells 2–4 µm in diameter, which are probably maturing spermatozoa, or small filaments, 4 µm long by 0.5 µm in diameter, which are probably sperm cells. An envelope of elongate cells is visible around some of these presumed sperm cysts, which are devoid of spicules.
Distribution and Ecology. All specimens were collected by trawl or dredge from 758–952 m and 1009–1019 m depth, SE of New Caledonia between 23°18 S 167°57 E and 23°23 S 167°54 E. The enlarged, solid attachment base of some of them indicates that they were living on rocky substrate.
Etymology. microstrongylata , for the numerous, characteristic microstrongyles of the species.
Remarks. This species has an external shape often found in Cladorhizidae and well documented in Abyssocladia . In this genus, the new species is well characterized by the presence in great number of microstrongyles forming a dense coating around the main axis and the filaments. This type of spicule has not been reported in Abyssocladia nor more generally in Cladorhizidae . Strongyles of rather similar shape are found in A. boletiphora Hestetun, Rapp & Xavier, 2017 , but they are larger and do not constitute a thick coating around the axis and the filaments.
The spicule slides made from diverse fragments show a certain variability especially in microscleres, possibly related to the presence of embryos which may contain spicules not found in the skeleton of the adult sponge.
The presence of such a remarkably dense coating of microstrongyles on the surface of the sponge raises the question of the role such a cover plays in prey capture. This will be discussed in a general remark below.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |