
Minutella minuta (Cooper, 1981)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3746.1.7 |
|
publication LSID |
lsid:zoobank.org:pub:40D305DB-2A50-476F-B7D9-B51673F4BAF9 |
|
DOI |
https://doi.org/10.5281/zenodo.6158522 |
|
persistent identifier |
https://treatment.plazi.org/id/FC5087EA-FFAE-FFF3-FF64-35C015E57811 |
|
treatment provided by |
Plazi |
|
scientific name |
Minutella minuta (Cooper, 1981) |
| status |
|
Minutella minuta (Cooper, 1981) View in CoL
( Fig. 2 A–O)
1981 Thecidellina minuta Cooper - p. 61, pl. 6, figs 27–40.
2009 Thecidellina minuta Cooper - Bitner, p.18, fig. 13A–I.
2010 Thecidellina minuta Cooper - Bitner, p. 653, fig. 5A–F.
2010 Minutella minuta Cooper - Hoffmann and Lüter, p. 148, pl. 2, figs 13–18; pl. 3, figs 13–15. 2013 Minutella minuta Cooper - Logan and Baker, p. 438, figs 1F, 4I.
Type locality. Samper Bank, south-east of Madagascar, western Indian Ocean, depth 380m (Cooper 1981).
Material. 28 specimens from two localities in the Sudan: 12 from GZ 93/8, 16 from SM 9B, including a total of 18 dorsal valves at both localities.
Diagnosis. (see Hoffmann and Lüter 2010, p. 148).
Description. Shell very small, mean length about 2mm, mean width about 1.5mm, longer than wide, biconvex, ventral valve larger than dorsal valve, cemented to substrate by cicatrix of attachment, anterior commissure rectimarginate, shell coarsely endopunctate, interarea of ventral valve with horizontal growth striations, not endopunctate, with central upraised elongate triangular rugideltidium with growth lines but without horizontal striations or endopunctae, lateral edge zones of rugideltidium feathered with underlying interarea. Ventral valve interior with prominent cyrtomatodont teeth covered with fibrous secondary shell, oval lateral adductor muscle scars, hemispondylium flat or slightly concave with no supporting septum, attached to floor of valve, with parallel prominent prongs apically pointed and slightly hooked at ends, interior of valve without prominent gonadal pits, interior granulose, margin with 3–4 rows of tubercles of fibrous secondary shell. Dorsal valve rounded, interior profile arched from anterior to posterior with median septum supported by thin plate spanning cavity in early growth stages ( Fig. 2 D), later filling in, median septum narrow, widening anteriorly, with central groove, margins slightly dentate, undamaged brachial cavities covered with reticulated canopy of spicules of variable arrangement (cf. Figs. 2 I and L) with a pair of ovate marsupial openings through canopy on either side of posterior end of median septum, bridge broad, no marsupial notch, median adductor muscle scars beneath bridge, with narrow delicate calcitic pole suspended from centre of bridge, no lateral outgrowths and not fused to cardinal process; cardinal process large, bilobate, lateral lobes with secondary shell covering, dental sockets flanking cardinal process, strongly tuberculated outer margins, with tubercles of secondary shell. Lophophore schizolophous.
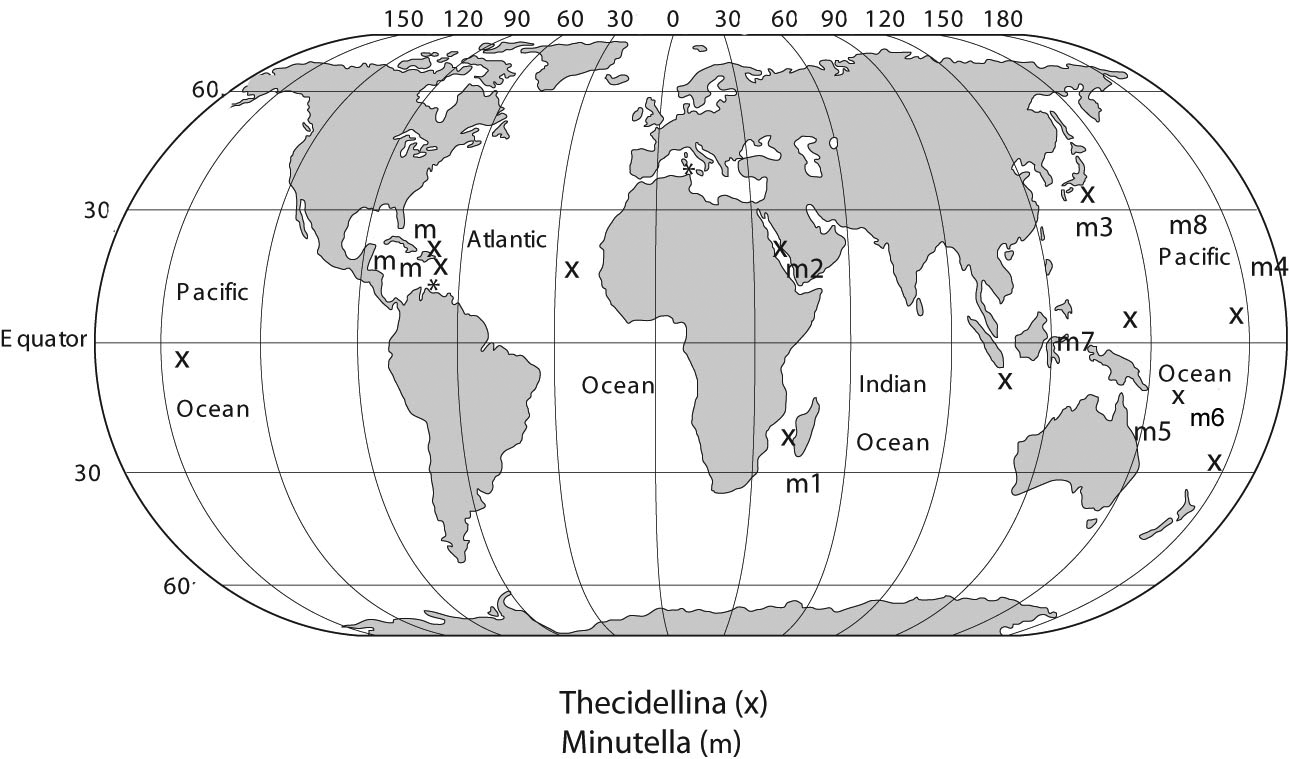
Remarks. Cooper (1981) described Thecidellina minuta from Samper Bank, south-east of Madagascar mainly on the basis of its small size, as the name implies. Hoffmann and Lüter (2010) included this species in their new genus Minutella for small thecidellinids with a pseudodeltidium (renamed rugideltidium by Logan and Baker 2013). The latter feature was not described by Cooper but its presence in the type specimens was confirmed by Hoffmann and Lüter (2010). These authors described M. tristani and M. bruntoni from the Caribbean and also recognized an Indo-Pacific group (or clade) consisting of M. minuta from Samper Bank, T. minuta from New Caledonia (Bitner 2009, 2010), and M. cf. minuta from Okinawa, Fiji, and Lizard Island ( Australia). To this list can be added Minutella cf. minuta from Sulawesi, Indonesia (Simon and Hoffmann 2013) and the Red Sea occurrences described here, as well as additional discoveries of this form from Palau and Saipan (our unpublished observations). This revised Indo-Pacific group is shown in Fig. 3 View FIGURE 3 as localities M1–9 and represents varieties of M. minuta with only minor differences insufficient to separate them as distinct species. The Red Sea specimens thus become the westernmost occurrence of a postulated Minutella minuta species complex stretching from the Red Sea to the central Pacific.
According to Hoffmann and Lüter (2010, p. 158, pl. 3) one of the main distinguishing characters separating the 2 groups is the nature of the dorsal valve median septum which is narrow and straight in the Indo-Pacific group, but broad and heavily tapering posteriorly in the Caribbean group. However, Red Sea specimens demonstrate that this feature can be quite variable (compare figures 2F, I, and L) and should be used with caution. Another difference is the presence in M. bruntoni and M. tristani of prominent median adductor muscle scars below the brachial bridge which are not seen in M. minuta .
Although there are 28 specimens of M. minuta from Red Sea sediments they are all devoid of soft parts. The living sites have not been found, although it is presumed that they are from cryptic habitats in nearby reefal environments (Zuschin and Mayrhofer 2009), perhaps similar to those for Minutella bruntoni from Grand Cayman which, with Thecidellina barretti (Davidson) and Argyrotheca woodwardiana (Davidson) , is common in low-light areas of reef caves (Logan 1983a). The Red Sea specimens often show abrasion or damage to the shell, suggesting post-mortem transportation from their living sites. The species has only been found at 2 localities (SM 9B and GZ 93/8) but its small size may have caused it to be overlooked elsewhere.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |