
Oribatula tibialis ( Nicolet, 1855 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.208456 |
|
DOI |
https://doi.org/10.5281/zenodo.5616825 |
|
persistent identifier |
https://treatment.plazi.org/id/FA7587D7-CC34-FFAA-FF55-620DFB10FA7E |
|
treatment provided by |
Plazi |
|
scientific name |
Oribatula tibialis ( Nicolet, 1855 ) |
| status |
|
Oribatula tibialis ( Nicolet, 1855) View in CoL
( Figs. 4 View FIGURE 4 E, 10D, 15–20)
Oribatula tibialis: Willmann 1931 View in CoL ; Schuster 1960; Wallwork 1983; Wunderle et al. 1990; Pérez-Íñigo 1993; Subías 2004, 2011; Weigmann 2006; Norton and Behan-Pelletier 2009. Notaspis tibialis Nicolet, 1855 .
Oribatula venusta Berlese, 1908 View in CoL .
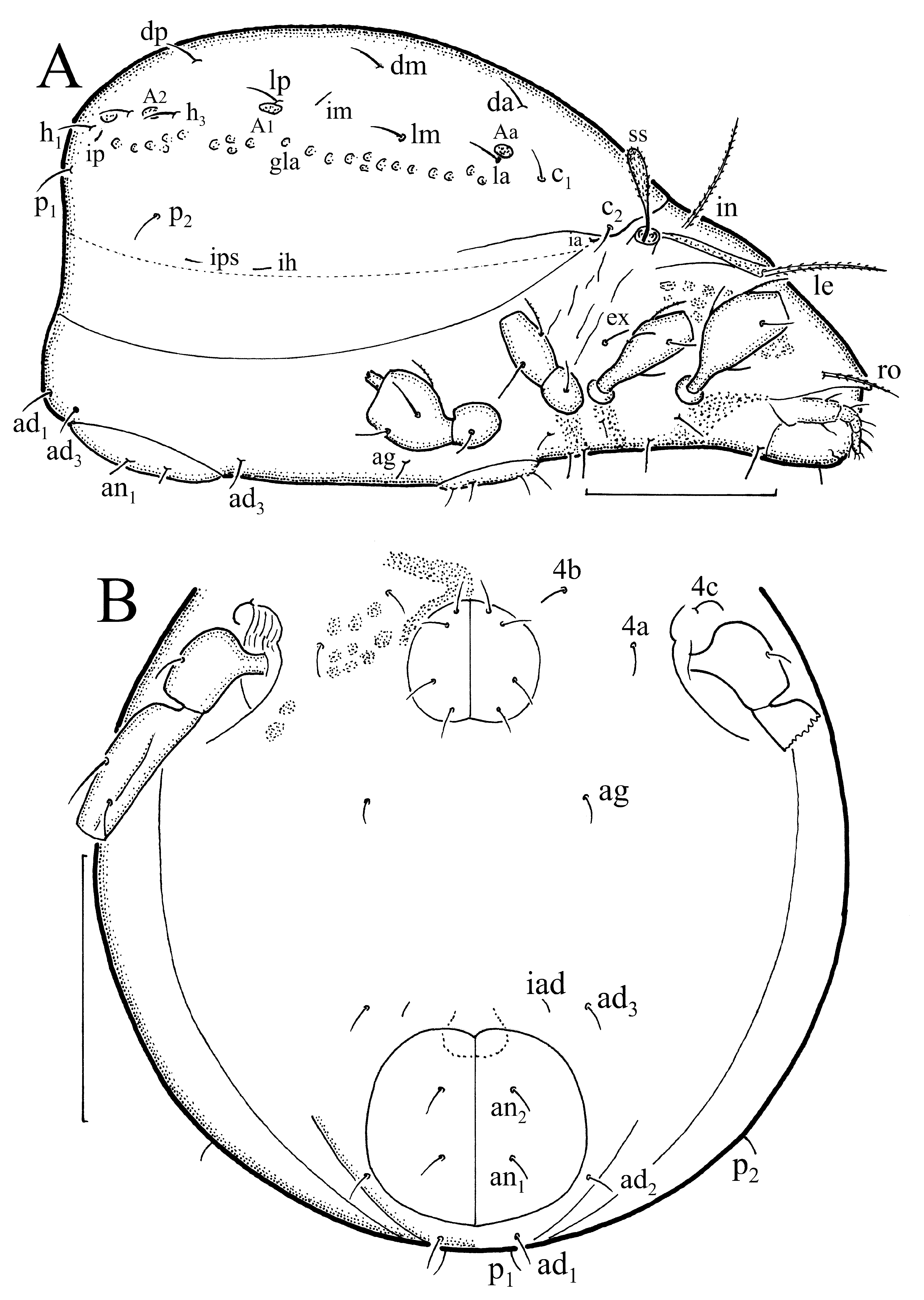
Diagnosis. Adult of medium size (410–530 µm), drop-shaped ( Fig. 15 View FIGURE 15 ), brown. Males usually smaller than females. Prodorsal seta le rather long, setae ro and in shorter; all barbed. Lamellae well developed, wider in anterior part than in posterior part, translamella absent. Sensillus rather short, clavate, with barbed head. Notogaster arched in lateral aspect ( Fig. 16 View FIGURE 16 A), with 13 pairs of short setae, and with rounded porose areas. Porose areas also on segments of legs, like tarsus I and II, tibia I–IV, and femur I–IV. Four pairs of genital setae, two pairs in anterior part, and two pairs in posterior part of genital plates ( Fig. 16 View FIGURE 16 B). Setae of ad -series small, ad 3 lateral to iad, other pairs posterior to anal opening. Solenidia and setae on tibia I and tarsus I similar ( Fig. 4 View FIGURE 4 E) as in Ph. lucorum , but porose areas on legs smaller. Formulae of leg setae (and solenidia): I – 1-5 -3(1)-4(2)-20(2); II – 1-5 -2(1)-4(2)- 16(2); III – 2-3 -1(1)-3(1)-15; III – 1-2 -2-3(1)-12. Tarsi tridactylous.
Juveniles oval in dorsal aspect, light-yellow, in some individuals with darker region gla. Larva with rather short setae, except longer prodorsal seta ro, nymphs with long and barbed prodorsal setae, except short seta ex, and rather short gastronotal setae, except longer setae c 3 and c 1. Sensillus short, clavate, with barbed head. Gastronotum arched in lateral aspect, with 11 pairs of setae in larva, and with 15 pairs in nymphs. Some gastronotal setae with excentrosclerites (c 2, la, lp and h 1 in larva, c 2, la, lp, h -series and p 1 in nymphs).
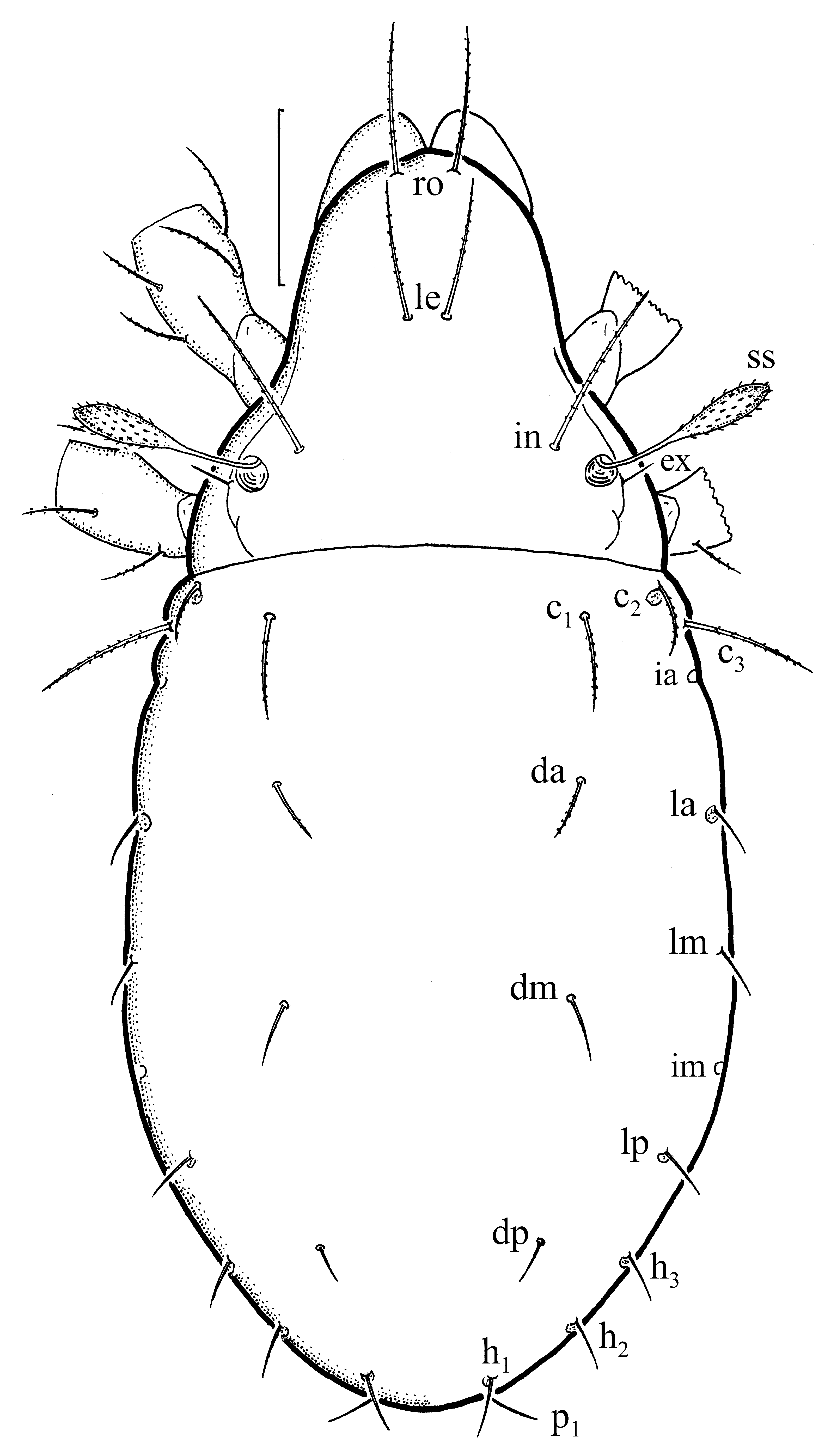
Description of juvenile stages. Larva oval ( Fig. 17), light-yellow, in some individuals with darker region gla. Prodorsum subtriangular, seta ro rather long, setae le and in shorter, and inserted wider; all setae slightly curved and barbed; seta ex short and smooth. Bothridium rounded, sensillus rather short, clavate, with barbed head.
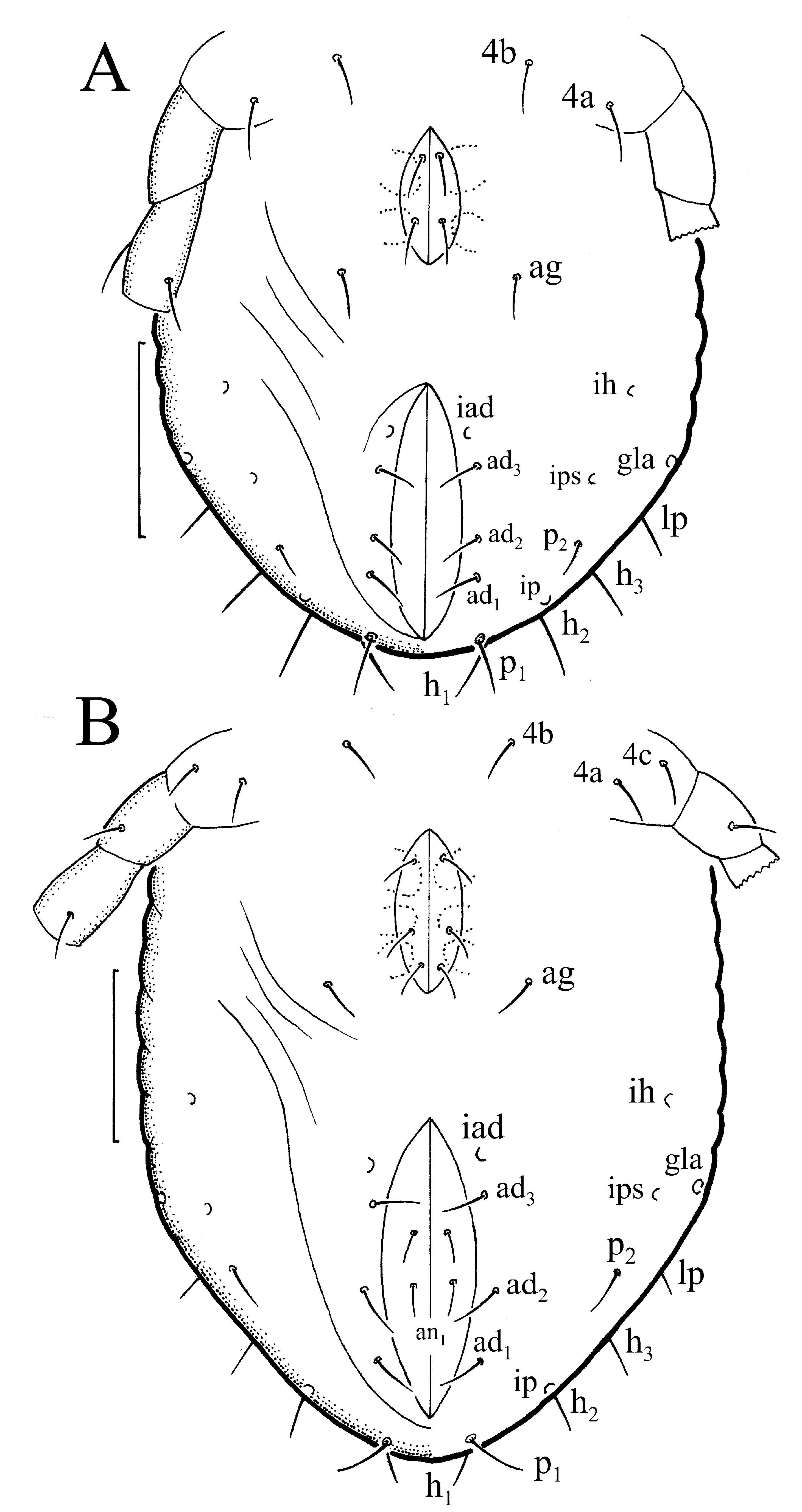
Gastronotum with 11 pairs of setae, including seta h 2, inserted lateral to posterior part of anal opening ( Fig. 18 View FIGURE 18 A); h 3 absent. All gastronotal setae rather short ( Table 1), except for slightly longer setae c 3 and h 1; all setae barbed. Setae c 2, la, lp and h 1 with excentrosclerites, other setae without excentrosclerites. Cupule ia posterior to seta c 3, cupule im posterior to seta lm, cupule ip anterior to seta h 1, cupule ih lateral to anterior part of anal opening. Paraproctal valves (segment PS) with two pairs of small setae. Anal region with few longitudinal folds.
Nymphs light-yellow and with relatively shorter prodorsum than in larva; some individuals with darker region gla. Protonymph with 14 pairs of setae, as seta h 3 and two pairs of p -series appear first time in this stage ( Fig. 18 View FIGURE 18 B) and remain in deuto- and tritonymph ( Figs. 19 View FIGURE 19 A, B). In deutonymph small setae of ad -series (ad 1– a d3) and pair ag appear and remain in tritonymph. Ontogeny of genital setae similar as in Ph. lucorum . Paraproctal valves of proto- and deutonymph glabrous, those of tritonymph with two pairs of small setae.
Prodorsal setae of tritonymph ( Fig. 20 View FIGURE 20 ) relatively longer than in larva, especially setae le and in; in longer than ro and le; all setae slightly curved and barbed, seta ex small and smooth. Bothridium rounded, sensillus clavate, with barbed head, slimmer than in larva.
Gastronotal setae rather short ( Table 1) and slightly curved, except longer setae c 1 and c 3; setae of c -series and da barbed, other setae rather smooth. Length of setae of d -series decreasing from anterior to posterior. Setae c 2, la, lp, h -series and p 1 with excentrosclerites, other setae without excentrosclerites. Cupules ia and im as in larva, cupule ip ventral to seta h 2, cupule iad lateral to anterior part of anal opening, cupule ips pushing lateral, and cupule ih pushing anterolateral to cupule iad ( Fig. 19 View FIGURE 19 B). Gland opening gla lateroventral to seta lp. Anogenital region with few longitudinal folds. Setation of tibia I and tarsus I of tritonymph ( Fig. 10 View FIGURE 10 D) similar to that of Ph. lucorum .
Summary of ontogenetic transformations. In the larva seta ro is longer than setae le and in, in the nymphs seta in is longer than setae ro and le, while in the adult seta le is longer than setae ro and in. The bothridium is rounded in all instars and the sensillus has clavate head, which is more slender in the nymphs and adult than in the larva.
The larva has 11 pairs of gastronotal setae (h 3 absent), the nymphs have 14 pairs (gained h 3, p 1 and p 2), while the adult loses seta c 3, and 13 pairs of setae remain. Lyrifissure ia is located as in Ph. lucorum . The formula of gastronotal setae in O. tibialis is 11-14-14-14 -13 (larva to adult), that of segments PS −AN is 22222-0333-022. The formulae of coxisternal, genital and aggenital setae are similar as in Ph. lucorum ( Table 2 View TABLE 2 ), all formulae are consistent with those described by Grandjean (1949).
Distribution and ecology. Oribatula tibialis is a Holarctic species ( Subías 2004, 2011; Weigmann 2006), which is considered to be eurytopic, but prefers forest ecosystems ( Schuster 1960; Wallwork 1983; Cianciolo & Norton 2006). It is also considered to be a pioneer in a succession of microarthropods in decomposing birch leaves ( Hågvar & Kjøndal 1981) and in primary succession of soil mites in a Norwegian glacier foreland ( Hågvar et al. 2009). According to Wunderle et al. (1990) it regularly colonizes the litter, but sporadically occurs in moss clumps and on tree trunks. Oribatula tibialis was also found in bird feathers ( Krivolutsky & Lebedeva 2002) and in the nests of seabird ( Coulson et al. 2009), like black-legged kittiwakes ( Rissa tridactyla L.), common eider ( Somateria mollissima L.) and glaucous gull ( Larus hyperboreus Gunnerus ) in Kongsfjorden ( Spitsbergen, Norway). In populations of O. tibialis females predominated ( Domes et al. 2007). It is an intermediate host of Moniezia expansa and M. benedeni ( Svadzhyan 1962) .
Oribatula tibialis View in CoL is considered to be microphytophagous ( Schuster 1960), and pollenivorous ( Wallwork 1983). According to Schneider and Maraun (2005) it prefers dark pigmented fungi (Codinea sp., Ulocladium sp. and Aureobasidium sp.) and belongs to fungivorous grazers ( Zaitsev et al. 2002).
In few papers the age structure of O. tibialis View in CoL have been investigated. In Ronda environs (Málaga, Spain) this species was more abundant in cypress litter than in pine and larch litter, and in summer the juveniles were distinctly more abundant than the adults ( Seniczak & Seniczak 2010c). In Korčula Island ( Croatia) it was also abundant in mosses, but the juveniles only slightly dominated the adults (Seniczak et al. 2011b). In Vilamorell near Borrassà (Cataluna, Spain, N42 o13.19, E2 o55.36, 78 m a. s. l) O. tibialis View in CoL dominated the oribatid mite community in broom ( Sarothamnus sp.) litter, reaching density of 325 individuals per 500 cm 3, and dominance index D = 69.4, with a large fraction of juveniles (67.7% of population).
The age structure of O. tibialis View in CoL was also investigated in polluted soils. This species tolerated copper smelter pollution ( Seniczak et al. 1997), being abundant in highly polluted soil, with a large fraction of juveniles (85% of population). However, it was sensitive to alkaline pollution ( Seniczak et al. 1999), and in the soil highly polluted by calcium it was not abundant and with smaller fraction of juveniles (37% of population).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Oribatula tibialis ( Nicolet, 1855 )
| Seniczak, Stanisław & Seniczak, Anna 2012 |
Oribatula tibialis:
| Willmann 1931 |
Oribatula venusta
| Berlese 1908 |
Notaspis tibialis
| Nicolet 1855 |