
Archaeonycteris relicta, Harrison & Hooker, 2010
|
publication ID |
https://doi.org/ 10.3161/150811010X504554 |
|
persistent identifier |
https://treatment.plazi.org/id/F84F2431-FFC5-A83E-A9B9-FE68FC019BA7 |
|
treatment provided by |
Valdenar |
|
scientific name |
Archaeonycteris relicta |
| status |
sp. nov. |
Archaeonycteris relicta sp. nov.
vidimus. (= v.) 1972.?Omomyoidea indet. Hooker: 180–181. v. 1980. Chiroptera View in CoL indet. Hooker & Insole: 38.
v. 1986. Microchiroptera gen. et sp. indet. 1 Hooker: 241, fig. 16.
Holotype
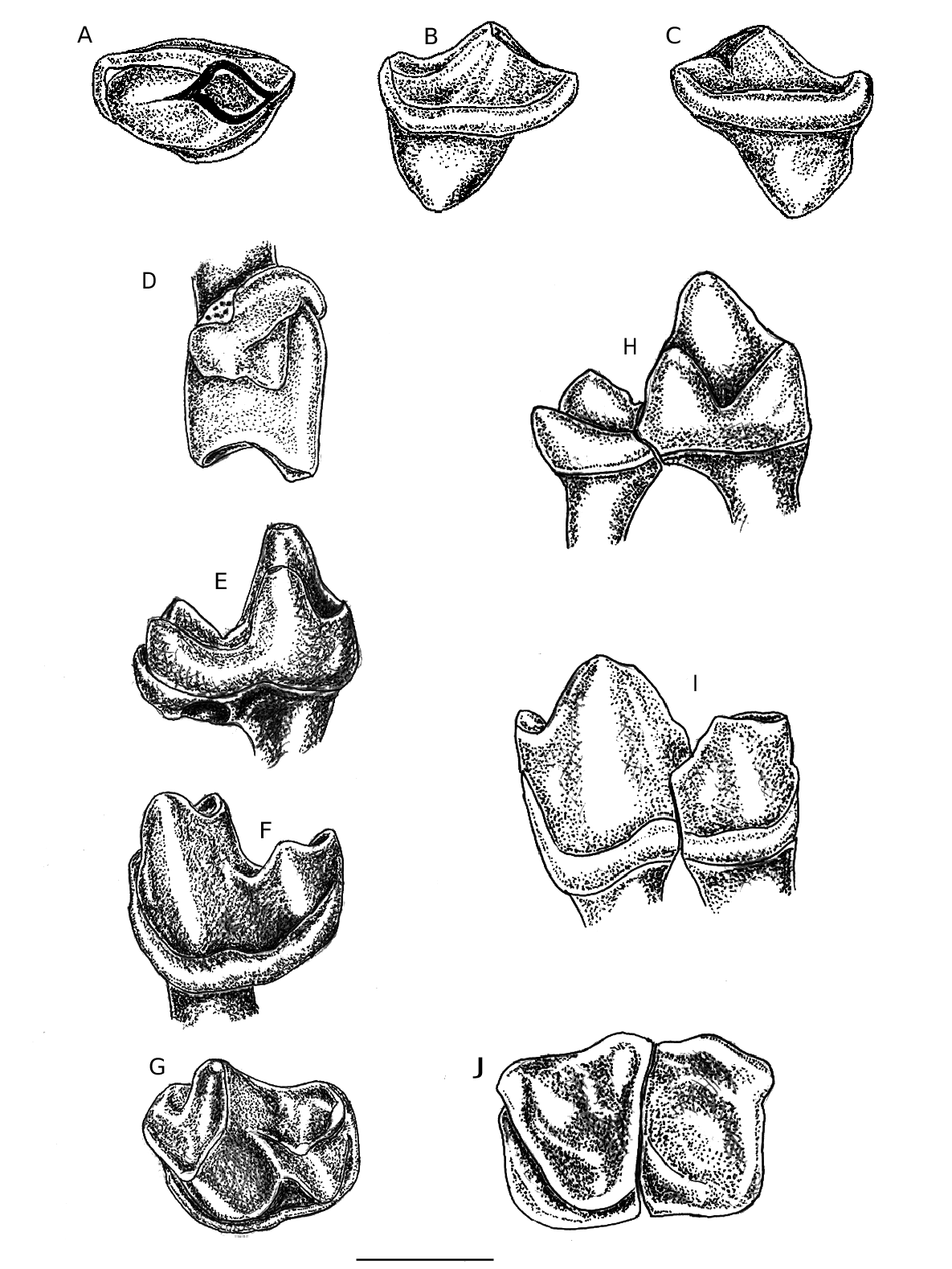
HZM 5.38291 View Materials isolated left M 3. CL 1.54; TRI W 1.12; TAL W 0.83 ( Figs . 1 View FIG D–G, 6D–F).
Paratype material
HZM 1.31799 View Materials right M 1/ 2 in maxillary fragment, metastyle broken CL–CW (protocone to parastyle) 2.34. Protocone lobe (a-p) 1.22 ( Figs. 2 View FIG A–D, 6J). HZM 7.38324 View Materials left M 1 View Left M /2. CL–CW (protocone to parastyle) 2.50 ( Figs. 2 View FIG E–H, 6K–L). HZM 4.37495 View Materials right P 2. CL 0.70 ; CW 0.45 mm ( Figs. 1 View FIG A–C, 6G–I). BMNH M 35710/12 Trigonid and talonid left M 1 View Left M /2. CL 1.98 (e) ; TRI W1.41; TAL W 1.31 ( Figs. 1 View FIG H–J, 6A–C). e = estimated measurement.
Doubtfully referred material from the type locality
HZM 16.31548 part protocone lobe right M 3. ( Fig. 2 View FIG I–J). HZM 3.35865 protocone lobe right M 1/2 ( Fig. 2 View FIG K–L).
Type locality
Creechbarrow, Dorset, Nat. Grid Ref. SY8240 9215, southern England.
Stratigraphy
Creechbarrow Limestone Formation Bed 12 ( Hooker, 1986), Bartonian, late Middle Eocene, Robiacian ELMA.
Referred specimen
BMNH M 29090, right M 3 talonid ; bed F?, Barton Clay Formation, Bartonian, late Middle Eocene; Barton, Hampshire ( Hooker, 1986: 241–243, fig. 16f–i).
Etymology
Latin relicta : alludes to a survivor of an archaic lineage.
Diagnosis
A small Archaeonycteris , mesial CW of M 1/2 2.34–2.50, CL of M 3 1.54.
Protofossa of M 1/2 bucco-lingually short, the transverse width subequal with its mesio-distal dimension, instead of clearly exceeding it, as in the three previously described European species listed below. There is no post- or premetaconule crista. Lower molars M 1 –M 3 with the paraconid low, not prominent, and the talonid shorter than the trigonid and with the entoconid virtually absent, forming only a crestiform bulge lingually, lower than the hypoconulid.
Description
HZM 1.31799 is a typical Archaeonycteris right M 1/2 with no mesostyle, hypocone or conules. The crown is shorter transversely than in A. trigonodon , A. pollex and A. brailloni . The protofossa measured from the protocone cusp to the median point of the centrocrista is shorter than the mesio-distal dimension, instead of clearly exceeding it as in the other described species above. There is a wide and shallow, buccally convex U-shaped centrocrista. The postprotocrista is inserted at the base of the metacone, as in A. pollex , not connected to the metacingulum as in A. brailloni . The crown width of this M 1/2 is notably less than in A. brailloni (3.0– 3.1 mm — Russell et al., 1973: 34). The specimen has the metastyle broken away, but a wide shallow ectoflexus appears to have existed. The resulting ectoflexus is not as deep as in the M 1 of A. brailloni and which is more compatible with M 2 ( Russell et al., 1973: fig. 6). The tooth is in situ in a maxillary fragment, which has a small nutrient foramen penetrating the palate below the preprotocrista.
F H E
G
The primary cusps, protocone, paracone and metacone, are conical. The tip of the metacone is damaged mesio-buccally. No small crests emanate from the base of the paracone or metacone as in some specimens of A. brailloni ( Russell et al., 1973: 30) and there is no trace of any pre- or post-paraconule crista or of the paraconule or metaconule on the pre- and postprotocrista. The protofossa is short and deep. The preprotocrista passes mesial to the paracone towards the parastyle. There is no hypocone, but a distinct angularity at the disto-lingual base of the protocone.
The parastyle is massive, with a bluntly rounded tip projecting mesially. The apparent absence of any buccal cingulum or stylar cusps, which are well developed in A. brailloni could be due to enamel erosion evident on the buccal margin of the tooth. The metastyle is missing, but it is clear that the buccal edge was recessed, without any trace of a mesostyle.
The protocone is surrounded lingually by a wide cingulum, which terminates abruptly on the mesial border of the crown, just in front of the level of the protocone tip. Distally the cingulum forms a straight border to the protocone lobe and is angulated outwards at the base of the metacone, where it is interrupted by a crack in the specimen. There is no evident internal distortion of the metacone base resulting from the crack. The tooth was originally threerooted, of which only the robust mesio-buccal root remains intact.
HZM 7.38324 ( Figs. 2 View FIG E–H, 6K–L) is a left M 1 View Left M /2, lacking the metacone and metastyle, which are broken off. The protocone and paracone are low, tubercular cusps, virtually unworn. The strong preprotocrista extends via the paracingulum to the blunt, broad parastyle, with a rounded mesiobuccal angularity. The paracone is slightly lower than the protocone and the preparacrista ends buccally with a small stylocone. The protofossa is deep and like HZM 1.31799 it is relatively short transversely. Judging from the area of its broken base the missing metacone was smaller than the paracone and there is no hypocone. The protocone is slightly slanted mesially. The preserved part of the ectoflexus is mesially positioned. It is shallower than in A. brailloni .
A robust cingulum surrounds the protocone base. Distally it is broken off close to the metacone; mesially it extends buccally to join the paracingulum. The buccal surface of the paracone is concave. A broad buccal cingulum is present, broken off at the same level as the centrocrista. Three roots were originally present.
HZM 16.31548 ( Fig. 2 View FIG I–J) comprises the mesial half of the protocone of a right M 3 with a strong mesial cingulum, inserted below the preprotocrista in a similar position to HZM 1.31799. The protocone is slightly more curved buccally than in the latter specimen. The paracone has been obliterated. The specimen is considered to be part of an M 3.
HZM 3.35865 ( Fig. 2 View FIG K–L) is a fragment of the protocone of a right M 1/2. Apart from the strong cingulum only the high protocone cusp and part of the preprotocrista are preserved. It appears distinctly larger than HZM 1.31799, but has a very similar shallow hollow between the preprotocrista and the robust cingulum.
HZM 4.37495 is a right P 2 ( Figs. 1 View FIG A–C, 6G–I), resembling in most respects this element in A. brailloni ( Russell et al., 1973) . Although the mesially situated cusp tip is worn, it is clearly low-crowned, with its buccal border quite strongly convex and the lingual border almost straight. A mesial cristid curves lingually to the crown tip. A distal cristid is almost straight and is evidently situated more lingually than in A. brailloni . In this specimen the talonid is slightly narrower than in A. brailloni and distinctly upturned. A relatively very strong cingulum surrounds the crown, eroded distobuccally. The single robust root is angulated distally below the crown. It may be noted here that the lower premolars of Stehlinia are high crowned and sharply point- ed. Small single-rooted anterior lower premolars resembling this in shape are not present in hipposiderids or rhinolophids.
M35710/12 is a left M 1 View Left M /2 talonid and trigonid described and figured by Hooker (1986: 242, fig. 16a–e). Found in the same hole, they were judged to belong to the same specimen, although the fit is not perfect. The specimen, remeasured from a cast after repositioning of the two fragments, gives a revised CL 1.98; TRI W 1.41; TAL W 1.31 ( Figs. 1 View FIG H–J, 6A–C).
HZM 5.38291 holotype ( Figs. 1 View FIG D–G, 6D–F) is an intact left M 3. It resembles in most respects the M 1/2 M35710/12, but the more reduced talonid shows that it is an M 3. It is well preserved, apart from the broken paraconid and slight enamel loss at the distolingual base of the crown. Only the cusp tips of the protoconid and metaconid are lightly worn.
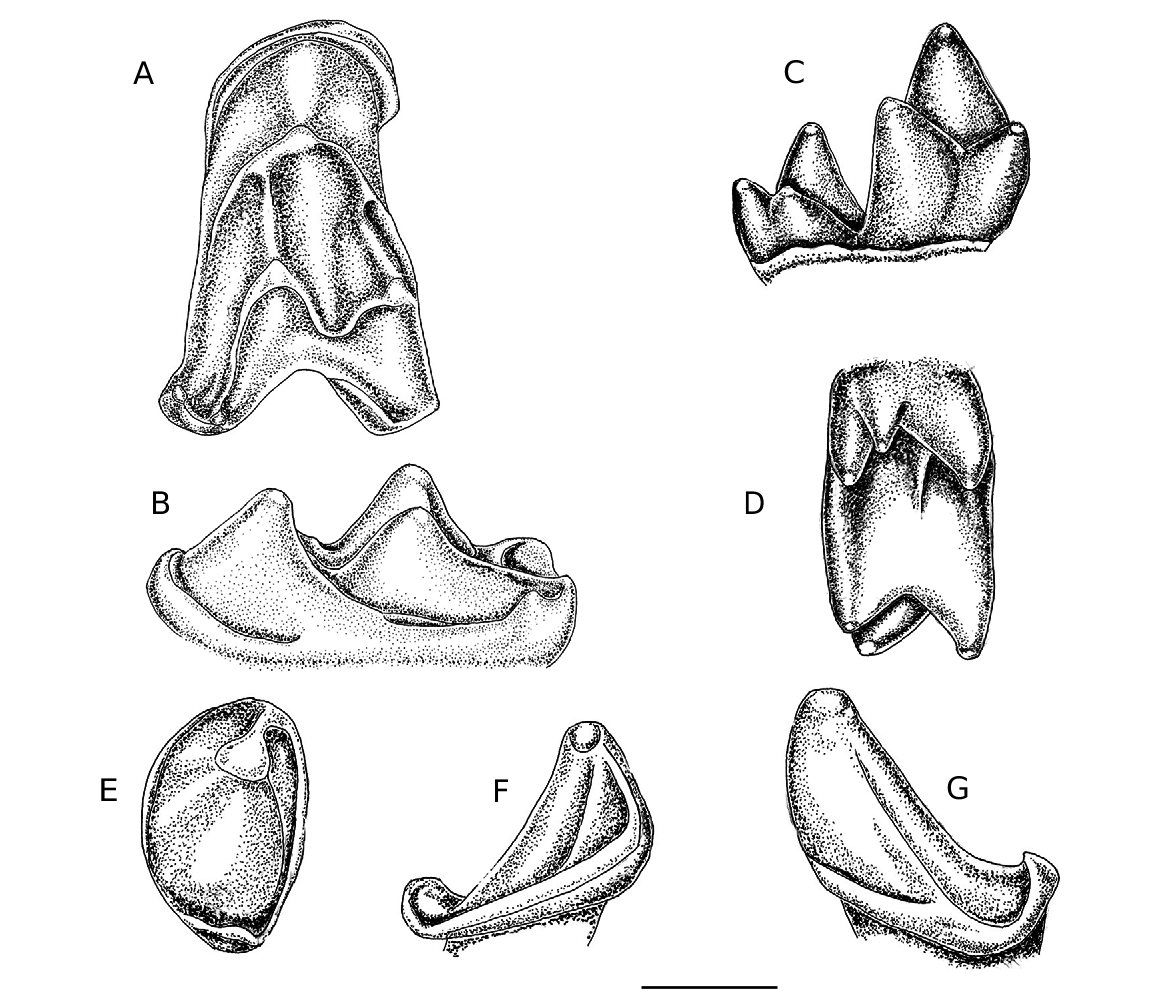
It is similarly a low crowned tooth, with the talonid distinctly shorter than the trigonid. The protoconid is dominant, the metaconid rather lower and the eroded paraconid appears to have been lower and less prominent than in the M 1/2 above. In the talonid the hypoconid is distinct, but low. The hypoconulid is almost central, projecting distally slightly less than the M 1/2. The entoconid is virtually absent, forming only a slight crescentic bulge disto-lingually. The talonid is distinctly more reduced than that of A. brailloni M 1/2 ( Fig. 3 View FIG C–D). A strong continuous buccal cingulum extends from the precingulid to the postcingulid. It is less angulat- ed below the protoconid than that of the M 1/2. No lingual cingulum is present. The short cristid obliqua is inserted almost centrally below the trigonid notch and it is notched near its junction with the trigonid. The mesial root of the tooth is preserved, the distal one has broken off.
Hooker (1986: 242, fig. 16f–i) described a right M 3 talonid (M29090) TAL W 0.93 from the Barton Clay (?bed F) sharing a number of features with the Creechbarrow M 1/2 and the M 3 described here. Comparison of this Barton Clay talonid with the
holotype M 3 above shows that it is identical in structure and it is here confidently referred to A. relicta .
Comparisons
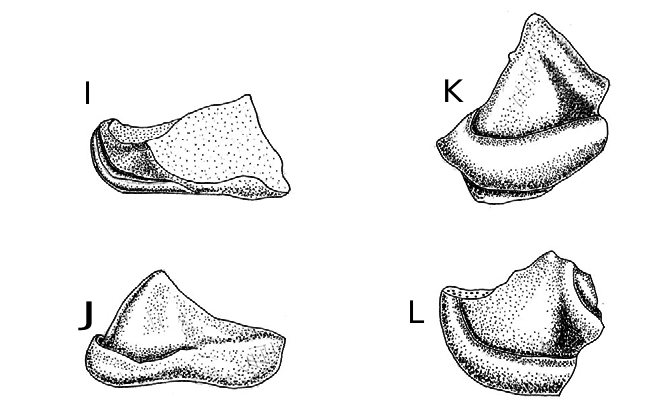
Most of the teeth of Archaeonycteris recovered are fragmentary. HZM 1.31799, the most complete M 1/2 so far found differs from previously described species in having a much smaller protofossa. This condition is linked with smaller size of the talonid of the lower molars and with the reduced entoconid. This nearly intact M 1/2 exhibits three of the diagnostic characters of Archaeonycteris ( Russell and Sigé, 1970: 125) . 1. Absence of the mesostyle. This is accompanied by a deep ectoflexus, listed by Simmons and Geisler (1998: 42) as the first of the apomorphies distinguishing Archaeonycteris . 2. Absence of the hypocone. 3. Absence of the metaconule, although a small paraconule may be present ( Russell and Sigé, 1970: 125). The conules are small or absent.
HZM 1.31799 (probably an M 2) and HZM 7.38324 are less transversely elongated than in all three European species, with a distinctly smaller protofossa and shallow and U-shaped centrocrista. The apparent absence of buccal cingulum and cingular cusps on the parastyle of HZM 1.37199 may be, at least in part, due to erosion of the enamel. This tooth resembles A. brailloni ( Fig. 3 View FIG A–B) in having an indication of a hypoconal lobe, absent in HZM 7.38324; this feature is not constant in A. brailloni ( Russell et al., 1973: fig. 6). It resembles A. pollex in having the post-protocrista directly inserted on the base of the metacone, rather than continuing into the metacingulum as in A. trigonodon and A. brailloni . The lower molars of A. relicta have reduced talonids, with the entoconid virtually absent.
Comparison is necessary with M 1/2 of some other genera of European Early and Middle Eocene Chiroptera and recently described Indian archaeonycterids.
Ageina tobieni Russell, Louis and Savage, 1973 is a taxon of uncertain familial status, most recently referred to Nataloidea incertae sedis ( Simmons and Geisler, 1998: 134, table 8). The M 1/2 show some similarity to the Creechbarrow M 1/2 (HZM 1.31799) in having a rather subquadrate basal outline, an ectoflexus, absent mesostyle and prominent para and metastyles ( Russell et al., 1973: fig. 8). It is clear, however, that this form differs in having a sharply angulated centrocrista. The trigon basin of the Creechbarrow M 1/2 is distinctly smaller and rounded buccally. The lingual cingulum is incomplete at the base of the protocone in Ageina tobieni , strong and complete in the Creechbarrow A. relicta . (see below for comparison of Ageina with an unidentified Microchiropteran trigonid).
Stehlinia Revilliod, 1919 . Bats of this genus, referable to the Nataloidea according to Simmons and Geisler (1998: 136, table 8) have an M 1/2 with a pronounced mesostyle and small crests originating from the bases of the paracone and metacone.
Icaronycteris ? menui Russell, Louis and Savage, 1973 has the M 1/2 more transversely elongate in form and also has the centrocrista sharply angulated. The lingual cingulum is variably complete ( Russell et al., 1973: fig. 4)
Palaeochiropteryx Revilliod, 1917 . Bats of this genus have a well developed mesostyle and the ectoflexus is shifted mesially with virtually absent concavity between the mesostyle and metastyle, as well as a sharply angulated centrocrista ( Russell and Sigé, 1970: figs. 4, 6 and 7, plate 1).
Protonycteris gunnelli Smith, Rana, Missaien, Rose, Sahni, Singh, and Singh, 2007 was recently described from the Ypresian of Vastan Lignite Mine, Gujarat, India ( Smith et al., 2007: 1004), based on the unique holotype left dentary, with P 3 –M 3 in situ. It is slightly larger than A. relicta (M 1 CL 2.00; TRI W 1.40; TAL W 1.40; M 2 CL 2.10; TRI W 1.50; TAL W 1.40; M 3 CL 1.90; TRI W 1.35; TAL W 1.05). It resembles A. relicta in having a reduced entoconid, but differs in having a distobuccally incomplete ectocingulum and short weak cristid obliqua in the lower molars. The talonid of M 3 is longer and the hypoconulid is shifted lingually. The maxillary dentition is as yet undescribed.
Archaeonycteris ? storchi described from the same Indian Early Eocene site ( Smith et al., 2007: 1007) and known from a left dentary fragment with M 1 –M 3 in situ is rather smaller in size than A. relicta (M 1 CL 1.50; TRI W 0.95; TAL W 1.00; M 2 CL 1.55; TRI W 1.05; TAL W 1.00). It has a more distinct and relatively stronger entoconid, the less reduced talonid thus differing strikingly from A. relicta . The maxillary dentition remains undescribed and its inclusion in Archaeonycteris remains uncertain.
Archaeonycteris ? praecursor Tabuce, Antunes and Sigé, 2009. This taxon has been recently described from a single right M 1/2 (UNLSNC — 447) from the Early Eocene of Silveirinha, Portugal. This tooth, with a CL 1.45 and CW 0.95 is slightly small- er than the two lower molars at present known of A. relicta and differs strikingly in morphology. Its cusps are contrastingly tall and pointed, with a more prominent, mesially directed paraconid. The entoconid is furthermore well developed, subequal in height with the hypoconid; the hypoconulid is more prominent and high, sublingual in position ( Tabuce et al., 2009). Whatever the ultimate generic status of this bat may prove to be, it is clearly quite different from A. relicta .
Rather few measurements are available for the lower molars of Archaeonycteris . CL of M 1 –M 2 2.40–2.48 (n = 4, m = 2.45); M 3 CL 2.12–2.20 (n = 2, m = 2.16) in A. pollex ; CL M 1 –M 2 2.00–2.20 (n = 4, m = 2.09); M 3 1.88 (n = 1) in A. trigonodon ( Storch and Habersetzer, 1988: table 2); A. brailloni M 1 CL 2.30 (n = 1). These measurements (crown widths not available) suggest that the lower molars of A. relicta are smaller, consistent with the reduced talonids observed. In A. pollex the relatively large hypoconulid is very high, as high as the entoconid or higher (Habersetzer and Storch, 1988). In A. brailloni the talonid cusps are all better developed than in A. relicta with the entoconid only somewhat reduced ( Fig. 3 View FIG C–D).
Discussion
This is the first confirmed occurrence of Archaeonycteris in the British fossil record. Savage and Russell (1983) recorded ‘ Archaeonycteris ’ from the Bartonian of Robiac but all other records of the genus are from older rocks. Legendre et al. (1992: 421, fig. 7a–b) figured an archaeonycterid of undetermined genus and species from Vielase, Phosphorites du Quercy, Early Eocene. Their figured upper molar (fig. 7a) has a transversely elongated trigon basin quite unlike A. relicta . The occlusal view of the figured M 1/2 shows an entoconid better developed than in A. relicta . Discovery of more intact material from the British Bartonian is needed to allow comparison of other tooth types.

| TRI |
TRI |
| H |
University of Helsinki |
| G |
Conservatoire et Jardin botaniques de la Ville de Genève |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Archaeonycteris relicta
| Harrison, David L. & Hooker, Jeremy J. 2010 |
Microchiroptera
| Dobson 1875 |
Chiroptera
| Blumenbach 1779 |