
Eudocima phalonia ( Linnaeus, 1763 )
|
publication ID |
https://doi.org/10.11646/zootaxa.5148.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:20BC627E-56A1-4674-A6B2-96F9B8DB15F7 |
|
DOI |
https://doi.org/10.5281/zenodo.7636003 |
|
persistent identifier |
https://treatment.plazi.org/id/ED434E3C-FFFD-FFC1-FF71-FA50FCA8F83B |
|
treatment provided by |
Plazi |
|
scientific name |
Eudocima phalonia ( Linnaeus, 1763 ) |
| status |
|
Eudocima phalonia ( Linnaeus, 1763)
( Figs 16 View FIGURE 16 :C–F, 21:K–L, 27:M–T, 28:A–B, 32:L, 33:C–D, K–L, 34:D–F, 35:D–F, 36:D–F, 37:B, G, 38:E–G, 39:E–G, 41:E, 43:E–F, 44:E–G, 45:F, 46:E–G, 47: E–G, 48:D–E, 49:B, 50:D–E, 51:B, 52:D–E, 53:B, 54:D–E, 55:B, 56:E–F, 57:A, 58:B, F, 59:C–D, 61:C–D, 62:C–D, 63:C-D, 64:C–D, 65:C–D, 66:C–D, 67:A, C, F, 68:D, 69:E–H, 71:B, 72: B, 87:B–E)
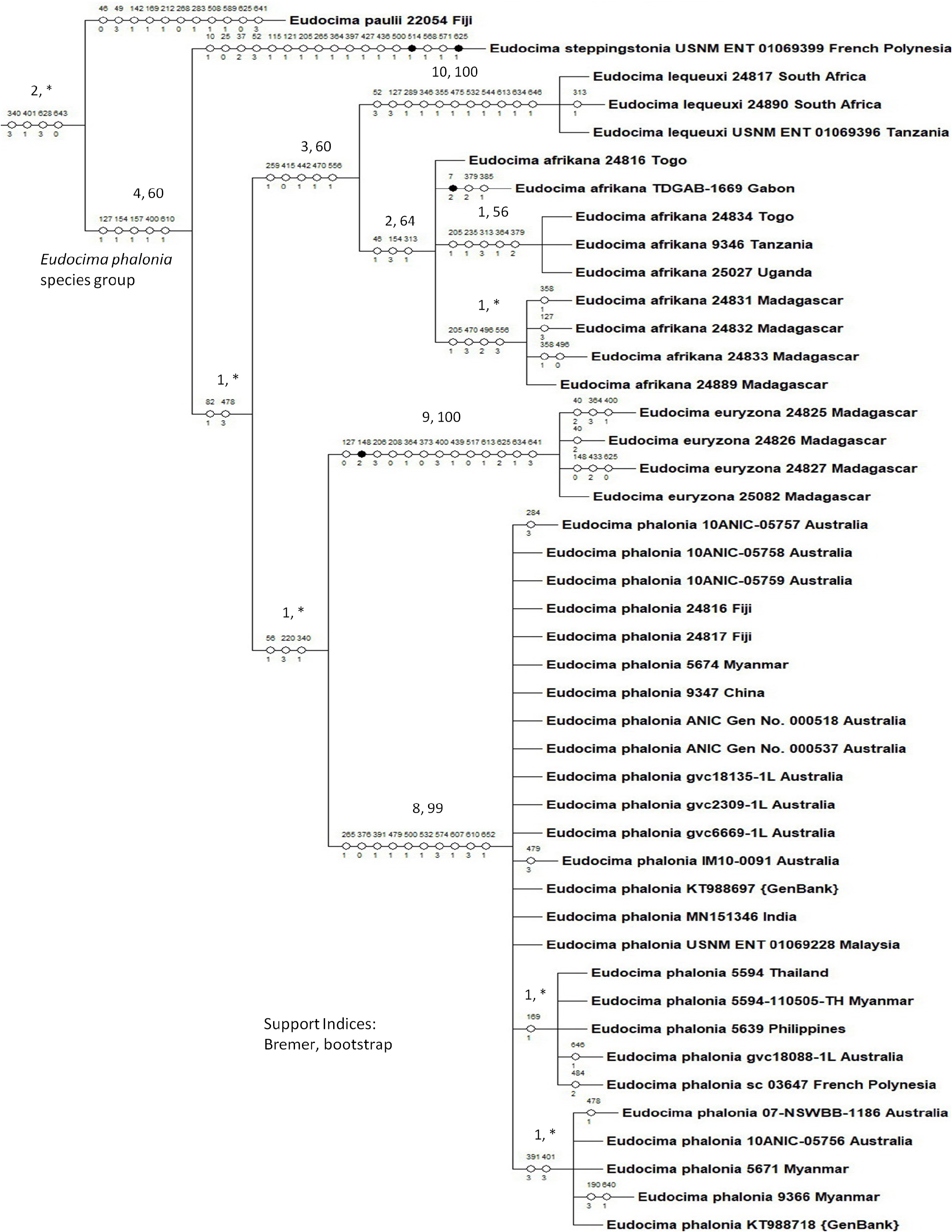
Diagnosis. Separation of this species from the allopatric E. afrikana , E. euryzona , and E. lequexui , and sympatric E. oliveri , is covered in the E. afrikana description.
Eudocima phalonia has also been confused with co-occurring E. homaena and E. cajeta . For example, Gilligan & Passoa (2014) is an often-cited reference for screening for E. phalonia ; however, their figure 2 of a photo of a live E. homaena from India is misidentified as E. phalonia . This same E. homaena photo is used again to represent E. phalonia on Wikipedia (2021). The third specimen in Gilligan & Passoa (2014) figure 4 is identified as a female E. phalonia , but it is actually a male. For E. homaena , only the males are likely to be confused as the females have a transverse green forewing band not present in E. phalonia . The males of E. phalonia have a fairly uniformly patterned forewing, whereas males of E. homaena have contrasting paler areas distal to the subterminal line and between the postmedial and antemedial lines. Also, E. phalonia has a contrasting lighter band on the posterior edge of the apical line, which may be greenish in fresh specimens. The male of E. homaena also has an apical line, but there is no contrasting lighter band or greenish coloration.
Males of E. phalonia and E. cajeta are somewhat similar in pattern, but in E. cajeta the apical line is faint if discernible, without the contrasting lighter band on the posterior side that occurs in E. phalonia . Also, E. cajeta has a more pronounced and pointed tornal hook relative to E. phalonia . For females, E. phalonia consistently has a larger whitish mark on the forewing than E. cajeta . Also, in E. phalonia the forewing has a glossier appearance with more contrasts relative to E. cajeta , which has a plainer, more granulated forewing.
Pattern of dimorphism in Fiji. In Fiji there are two male phenotypes of E. phalonia , the typical phenotype and a phenotype which is darker ventrally, particularly in the apical areas, with a reduced medial patch ventrally ( Fig. 21 View FIGURE 21 : K). Zilli et al. (2017) provided additional differences between these phenotypes, but reported an intermediate phenotype from Guadalcanal, and no meaningful differences in genitalia between the two phenotypes. We sequenced two of the darker phenotypes from Fiji (DNA sequence vouchers 24816 and 24817) and both have the most common COI 5’ haplotype found in typical E. phalonia ( Fig. 86 View FIGURE 86 ). We also dissected a male of one of these specimens (HLK: 2647) and found the genitalia to be the typical E. phalonia morphotype. Given the lack of differentiation in both male genitalia and COI 5’ plus the Zilli et al. (2017) report of an intermediate phenotype, we favor the hypothesis that the second Fiji phenotype represents infraspecific variation in E. phalonia .
Life history and bionomics. Most recent Eudocima life history studies have focused on E. phalonia . This species is associated with temperate broadleaf and mixed forests, tropical and subtropical grasslands, savannas and shrubs, and tropical and subtropical moist broadleaf forests ( Davis 2005). This species often exposes its bright hindwings while feeding, revealing the medial patch which is useful for separating it from other Eudocima species ( Fig. 87 View FIGURE 87 : B, E).
In a Sarawak orchard it had a 40-day life cycle from oviposition to adult emergence (Kueh 2012). The larvae of E. phalonia use Menispermaceae , but in the eastern part of their range they feed on Erythrina (Fabaceae) ( Cochereau 1977). The cocoon may stay on the host plant or drop with the leaves ( Waterhouse & Norris 1987). Bänziger (1982) noted that the three Tinospora (Menispermaceae) foodplants preferred in Thailand are most common in more open areas where these tenacious vines can endure drought and generate aerial roots. Foodplants thrive in close proximity to humans where they even survive on telephone posts and wires. In Palawan ( Philippines) E. phalonia occurred in disturbed forest but did not come to bait traps in primary forest openings (observation by RJB). RJB also didn’t find it in old growth forest in Papua New Guinea ( PNG) where it is widely distributed and not considered a pest. In PNG much of the forest remains intact and E. phalonia is believed to be regulated by native parasitoids such as Ooencyrtus sp. ( Hymenoptera : Encyrtidae ) and Telenomus lucullus Nixon ( Hymenoptera : Platygastridae ) which contributed up to 95% mortality of the eggs ( Sands & Liebregts 2005). Bänziger (1982) urged the establishment and protection of forest reserves for control of E. phalonia . He attributed the increase in E. phalonia to replacement of Tinaspora -poor primary forests with secondary forest habitat conducive to their larval foodplants, and to increased cultivation of fruit hosts for the adults.
Outbreaks of E. phalonia were reported by Cochereau (1977) in New Caledonia during February and March following a rainfall deficit of over 50% during September to December in the previous year. That outbreak caused an increase in damage to citrus production from 4% in a normal year to 90% after the drought period ( Cochereau 1973, 1977). Vargas-Fonseca et al. (2020) observe that the 1968–1969 drought in New Caledonia was prompted by the El Niño-Southern Oscillation (Benoir & Delcroix 2000) and that these El Niño related droughts likely also triggered other outbreaks of fruit-piercing moths. Intense rainfalls after drought periods can enhance vigorous new larval food production ( Ngampongsai et al. 2005). This young plant tissue with greater nitrogen and reduced defensive compounds ( Shure et al. 1998) can be linked to the improved survival rate of young larvae ( Leroy et al. 2021; Srygley et al. 2010, 2014).
Distribution and dispersal. This widespread species is found from India to Australia and east to Hawaii. It has become established on many isolated island chains in the southern Pacific. In Australia E. phalonia can move great distances between breeding sites and fruit orchards (Sands & Schotz 1988). In New Caledonia E. phalonia temporarily leaves the wild figs in the mountains to use larger orchard fruit on the plains but returns to mountain biotopes after fruits are harvested ( Leroy et al. 2021). The ability to migrate allows it to leave the tropics during hotter weather to temporarily colonize temperate regions in eastern Australia ( Sands et al. 1991). It was first recorded from Hawaii (Oahu) in 1985, and by 1986 it had spread to four other Hawaiian Islands (Kessing & Mau 1993). True E. phalonia does not occur in mainland Africa or Madagascar.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Calpinae |
|
Genus |