
Triturus ivanbureschi, Wielstra, B., Litvinchuk, S. N., Naumov, B., Tzankov, N. & Arntzen, J. W., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3682.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:69B9A846-616F-4774-B3F0-B796D2B90431 |
|
DOI |
https://doi.org/10.5281/zenodo.5658363 |
|
persistent identifier |
https://treatment.plazi.org/id/EB5087CA-FF96-5301-00DB-4BFCF7A9F6E1 |
|
treatment provided by |
Plazi |
|
scientific name |
Triturus ivanbureschi |
| status |
sp. nov. |
Triturus ivanbureschi View in CoL sp. nov. Arntzen & Wielstra
Type material. Holotype. RMNH. RENA.47200, an adult male from Ostar Kamak, Bulgaria (coordinates: 41.878°N, 25.853°E, elevation: 240 meter), collected 8 May 2012 by J.W. Arntzen, N. Tzankov and B. Wielstra ( Fig. 4 View FIGURE 4 ). Paratypes. Three males ( RMNH. RENA.47201-47203) and 15 females ( RMNH. RENA.47204-47218), with collection details identical to the holotype. For imagery, see online Appendix 5.
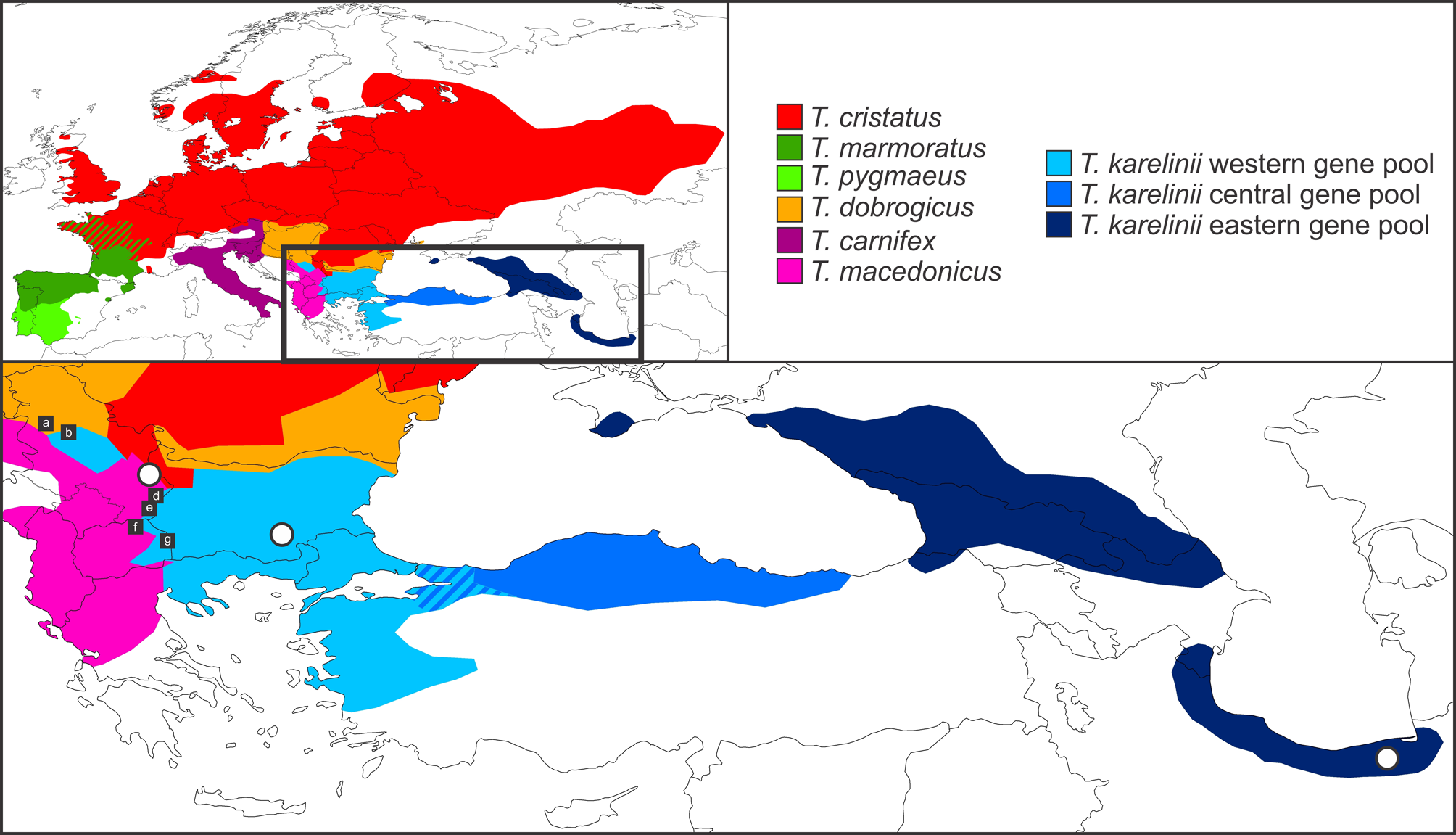
Diagnosis. The species is assigned to the genus Triturus because it possesses the characteristics of crested newts (the T. cristatus superspecies), in particular the combination of a denticulated crest, a dark brown dorsal coloration and orange ventral side covered with black spots. Based on mitochondrial DNA sequence data, crested newts from the type locality are firmly placed in the genus Triturus , within the western T. karelinii s. l. mitochondrial DNA lineage ( Wielstra et al., 2013b). As yet only molecular genetic data are available to distinguish T. ivanbureschi sp. nov. from its sister species T. karelinii . More details are provided in the section ‘A review of phylogenetic studies with Triturus karelinii s. l. ’. How to distinguish T. ivanbureschi sp. nov. + T. karelinii from the other crested newt species is discussed in the section ‘The name T. arntzeni is a junior synonym of T. macedonicus ’.
Description of holotype. A relatively robust crested newt, with a broad head (wider than neck) and well developed limbs. Four fingers and five toes, slightly fringed but interdigital webbing absent. Relative length of fingers 1 <4 <2 <3. Relative length of toes 1 <5 <2 <4 <3. Skin granulated on dorsal and lateral sides and throat; tail and venter smooth. Gular fold inconspicuous. Swollen cloaca with papillae bordering cloacal slit. Denticulated crest, interrupted over the position of the legs and cloaca. Laterally compressed tail with evident dorsal fin and less conspicuous ventral fin. In life a brownish-blackish base color on the dorsolateral side, with scattered black flecks, most pronounced and marbled on the head. Bluish-white streak along the lateral side of the tail along the caudal vertebrae. A bright-orange ventral side and throat, dotted with small angular black spots; throat spots smaller, denser and more angular than belly spots. In the preserved state colors have slightly faded ( Fig. 4 View FIGURE 4 ). The number of rib-bearing pre-sacral vertebrae (NRBV) is 14, as determined from X-ray photographs (see online Appendix 6). The following measurements are in millimeters and have been determined with plastic Vernier calipers. Snout-vent length up to and including the hind-legs 54.0; Snout-vent length up to and including the cloaca 61.0; Total length 106.5; Inter-limb distance 28.0; Head length 14.8; Head width 11.3; Length of the right arm including the third finger 21.9; Length of the right third finger 8.5; Length of the right leg including the fourth toe 21.4; Length of the right fourth toe 8.5.
Description of paratypes. The morphology of the paratypes resembles that of the holotype in general, but the pattern of black spots on the ventral side is highly variable among individuals (online Appendix 5). All paratypes have an NRBV count of 13, except RMNH.RENA.47205 (and probably RMNH.RENA.47202), which has an NRBV count of 14 (Table 2; online Appendix 6). Compared to males (such as the holotype), adult females have non-swollen cloacae, lack the denticulated crest and possess a less pronounced tail fin. In Table 2 the measurements taken for the holotype are provided for the paratypes.
Distribution. The European distribution encompasses the south-eastern Balkan Peninsula, covering most of Bulgaria, the eastern parts of Greece, Macedonia and Serbia, as well as European Turkey. An isolated distribution pocket is found in Serbia that is disconnected from the main range by other intervening Triturus species ( T. cristatus in the north and T. macedonicus in the south; Arntzen & Wallis, 1999; Wielstra & Arntzen, 2012). The Asian distribution is restricted to Asiatic Turkey, along the coast of the Aegean Sea, the Marmara Sea and the Black Sea, reaching up to c. 300 kilometers inland (usually less) but not inner Anatolia. An outline of the distribution is provided in Fig. 1 View FIGURE 1 ( T. ivanbureschi sp. nov. encompasses the western and central gene pools of T. karelinii s.l., which meet in northwestern Asiatic Turkey). An incomplete listing of locality data is provided in Wielstra et al. (2012).
Etymology. We dedicate the newly recognized species of crested newt to the memory of Dr. Ivan Buresch (1885-1980), scientific director of the Institute of Zoology, Sofia (see pp. 234-235 in Adler, 2012). Dr. Buresch, together with his assistant Jordan Zonkov, laid the foundation for herpetology in Bulgaria and indeed the Balkan region with the publication of the four part monograph, ‘ Untersuchungen über die Verbreitung der Reptilien und Amphibien in Bulgarien und auf der Balkanhalbinsel’ ( Buresch & Zonkow, 1933, 1934, 1941, 1942). The work is published in the Bulgarian language with extensive summaries in German. The compound name ivanbureschi was chosen to avoid confusion with ‘ Triturus cristatus karelinii forma bureschi ’ ( Wolterstorff, 1925) (nomen illegitimum, Litvinchuk et al., 1999; Mertens & Wermuth, 1960).
Proposed vernacular name. Balkan-Anatolian Crested Newt or Buresch’s Crested Newt.

| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.