
Melitaea timandra binaludica, Kolesnichenko & Kotlobay, 2022
|
publication ID |
https://doi.org/10.5852/ejt.2022.830.1865 |
|
publication LSID |
lsid:zoobank.org:pub:C1F47AD9-ECF6-4F0F-9928-55A45332FF4B |
|
DOI |
https://doi.org/10.5281/zenodo.6855860 |
|
persistent identifier |
https://treatment.plazi.org/id/62F42B37-68FD-4A67-8E4F-FBAA2ED93033 |
|
taxon LSID |
lsid:zoobank.org:act:62F42B37-68FD-4A67-8E4F-FBAA2ED93033 |
|
treatment provided by |
Felipe |
|
scientific name |
Melitaea timandra binaludica |
| status |
subsp. nov. |
Melitaea timandra binaludica subsp. nov.
urn:lsid:zoobank.org:act:
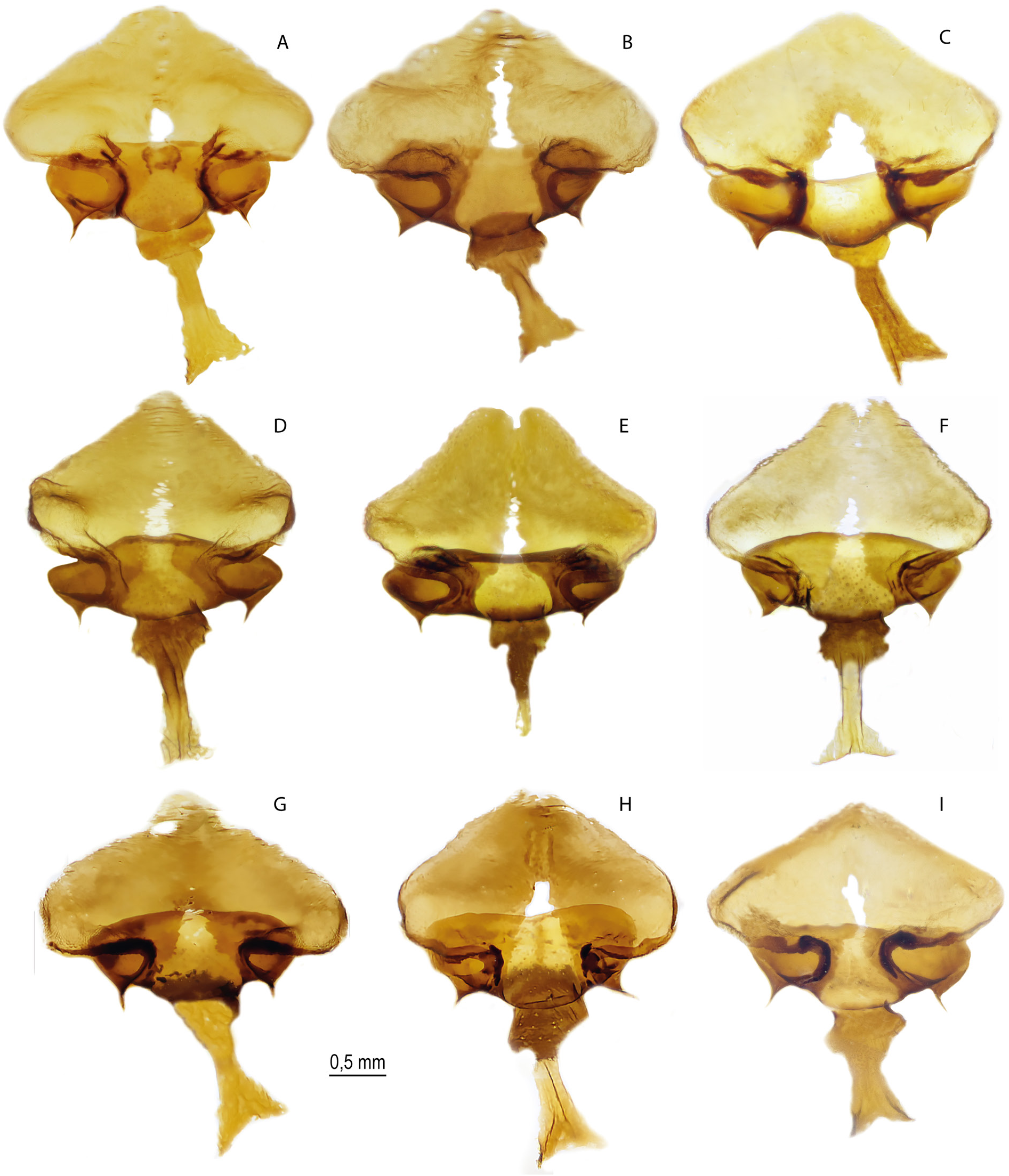
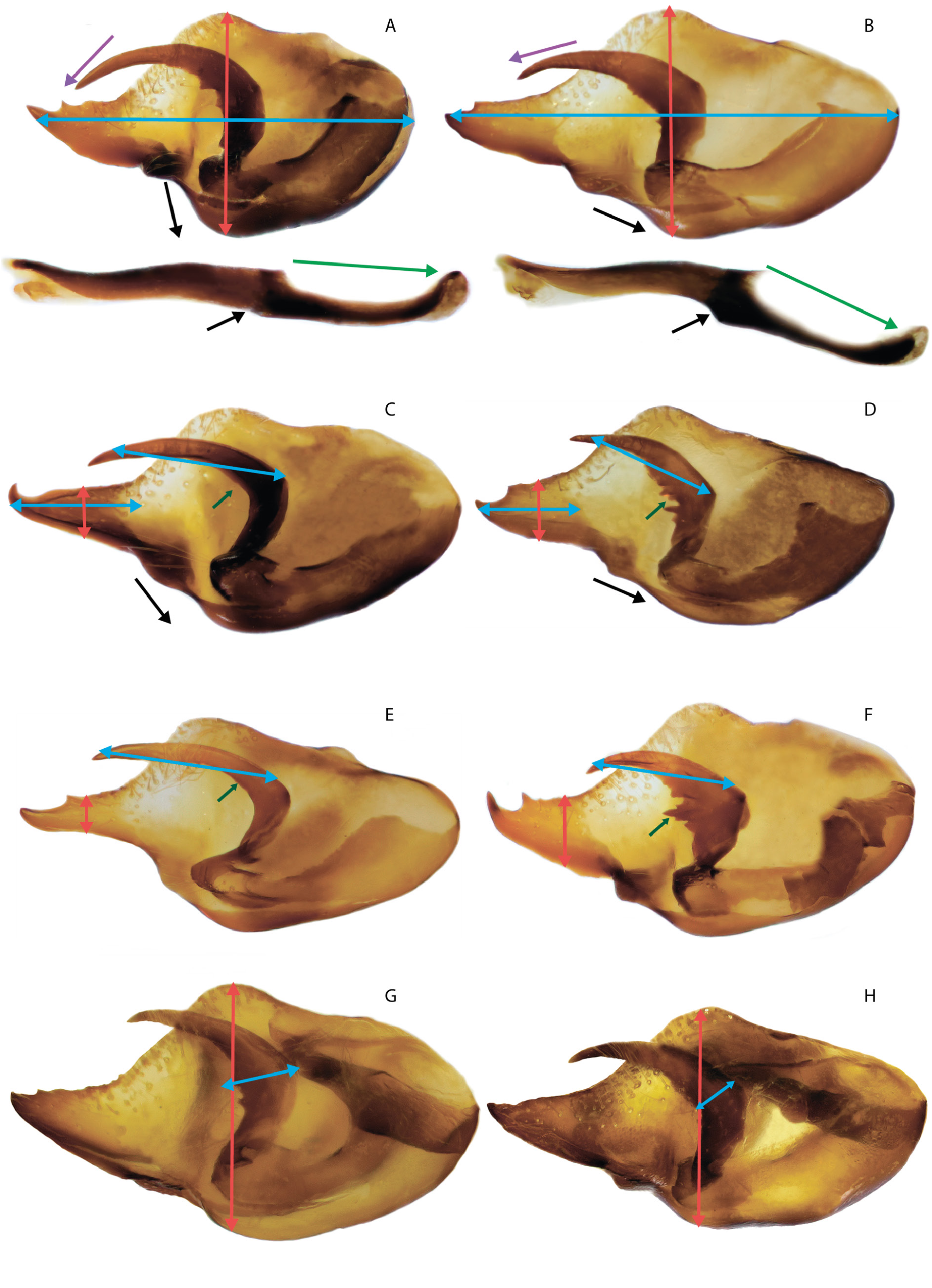
Figs 2B – D, F–H, J–L, N–P View Fig , 10G – O View Fig , 11D – I View Fig , 14–15 View Fig View Fig , 17D – I View Fig , 19D – I View Fig , 20 View Fig , 29B, D, H View Fig , 30 View Fig ; Tables 3 View Table 3 , 6
Differential diagnosis
Melitaea timandra binaludica subsp. nov. differs from the nominative subspecies in appearance by its smaller size, the absence or weakly expressed UPF postdiscal pale area in males, the presence of UPH discal row, the developed UPH basal suffusion and the presence of UNH dark scales both in males and females. In the structure of the male genitalia, the main distinguishing features should be considered a noticeably thinner caudal process of the valva (on average thinner than in the nominative subspecies) and a noticeably less wide central part of the harpe (1.5–2 times narrower than in the nominative subspecies). In addition, unlike the nominative subspecies, the harpe on its inner surface carries a smaller number of teeth.
Etymology
The name is toponymic and denotes the name of the mountain Ridge on which the type series was collected.
Type material
Holotype ( Fig. 15A, I View Fig ) IRAN • ♂; Rezavi Khorassan Prov., Kuh-e Binalud Mts, Dorrud v. vicinity ; alt. 2430 m; 12–13 May 2017; K. Kolesnichenko leg.; SDM .
Paratypes ( Fig. 15B– H, J–P View Fig )
IRAN – Rezavi Khorassan Prov. • 82 ♂♂ (7 dissected), 18 ♀♀ (6 dissected); Kuh-e Binalud Mts, Dorrud v. vicinity ; alt. 2430 m; 12–13 May 2017; K. Kolesnichenko leg.; EDMSU • 28 ♂♂ (20 dissected), 10 ♀♀ (7 dissected); Qadamgan area, Dorrud v. vicinity, Gerina v.; alt. 2000 m; 4 Jun. 2009; K. Kolesnichenko leg.; EDMSU • 2 ♂♂ (all dissected); Kuh-e-Binalud , 15 km SW of Zoshk; alt. 2300– 2500 m; 7 Jun. 1999; Eckweiler leg.; EDMSU • 11 ♂♂; 6 km N of Gerine, Kuh-e-Binalud Ridge ; 36°09′15.56″ N, 59°10′54.01″ E; alt. 2015 m; 9 May 2019; A.A. Kotlobay leg.; coll. A.A. Kotlobay. – Mazandaran Prov. GoogleMaps • 1 ♀ (dissected); S macroslopes of Albors Mts, 80 km SE of Sari, 5 km NE of Foulad Mahhaleh v., E slopes of Sultan Kuh Mt. ; alt. 2000 m; 2 May 2018; K. Kolesnichenko leg.; EDMSU • 1 ♀ (dissected); same locality as for preceding but 12 May 2019; A. Kotlobay leg.; EDMSU . – Horossan Prov. • 1 ♂ (dissected); 35 km N of Birjant t.; 28 Apr. 2006; K. Kolesnichenko leg.; EDMSU • 1 ♂ (dissected); 75 km N of Birjant t., Sedeh v. vicinity ; alt. 1500 m; 27 Apr. 2005; R. Nazarov leg.; EDMSU .
AFGHANISTAN – Bamian Prov. • 4 ♂♂ (all dissected), 1 ♀ (dissected); Band-e-Amir, Dzhudoi-Kvak Gorge ; alt. 3200 m; 1–2 Jul. 2009; O. Pak leg.; EDMSU • 1 ♂ (dissected); Hazarajat, Band-i-Amir ; alt. 9400–11000 ft; 9–10 Jun. 1960; Colin Wyatt leg.; EDMSU • 5 ♂♂ (all dissected), 1 ♀ (dissected); Band-e-Amir ; alt. 3200 m; 5 Jul. 2009; I. Pljushtch leg; EDMSU .
Description
Male ( Figs 2B–D, J–L View Fig , 15A–D, I–L View Fig )
WINGS. FW length is 16.5–18 mm, the holotype is 18 mm. UPS ground color is bright orange. UPS black pattern well developed; UPS thin black marginal border with well-defined proximal marginal spots along the outer edge of the wings; UPF submarginal row formed by black strokes, UPH – by thin lunules, usually not connected to the black marginal border; UPF discal row represented by well-defined black spots fused closer to the costa, UPH discal row is more or less developed and formed by small black spots reduced closer to the costa; UPF postdiscal pale-yellow area, located behind discal row, is weakly expressed or absent, usually there are 2–3 pale macules near the UPF costal edge; UPF paleyellowish macule in the distal part of the discoidal cell is usually absent; UPH discal row is formed by fine black spots; UPH basal suffusion covers up to ⅓ of the wing surface; UNF is bright orange with well-developed pale area along the outer edge of the wing between the veins Sc and M2, UNF pale macules in the postdiscal area is usually absent; UNH ground color is white, with an admixture of dark scales. UNH lunules forming the proximal edge of submarginal orange fascia outwardly concave and sharply pointed between veins M3 and Cu2.
MALE GENITALIA ( FigS 10G – O View Fig , 17D – I View Fig , 19D – I View Fig , 29B, D, H View Fig ). In general, the size of the genitalia is smaller than that of the nominative subspecies. Valva with a relatively narrow (on average narrower than that of the nominative subspecies) caudal process, usually with several small spines on the dorsal surface. The relatively short harpe is somewhat expanded in the central part and bears a small number of noticeable teeth on the inner surface. The expanded part of the harpe is 1.5–2 times narrower than that of the nominative subspecies. Aedeagus with a rounded convex dorsal edge, its posterior part is mostly located at an angle to the anterior part and is directed downward. When both parts of the aedeagus are joined, there is a well-defined protrusion on the ventral side. Narrow saccus is distally pointed, its width is 2–3 times less than the length.
Female ( Figs 2F–H, N–P View Fig ; Fig. 15E–H, M–P View Fig )
WINGS. FW length is 17.5–20 mm. Externally the female is similar to the male, but the pattern is more contrasting. UPF postdiscal pale area is more developed than in the male. There is a well-developed pale macule in the discoilal cell between the first and second discal spots. UPH submarginal row is usually reduced. UNS pattern is similar to that of males.
FEMALE GENITALIA ( Fig. 11D–I View Fig ). In general, the genitalia are similar to those of the nominative subspecies. The postvaginal plate is roundly triangular in shape. The antevaginal plate is narrow in the dorsoventral direction, its outer edge does not extend beyond the boundaries of the bend of the postvaginal plate (auricules).
Preimaginal stages: egg ( Fig. 20 View Fig , Table 3 View Table 3 )
Material examined: 4 specimens, 27 eggs, Iran, Rezavi Khorassan Prov., Qadamgan area, Dorrud v. vicinity, Gerina v., alt. 2000 m.
The description of the egg of M. timandra binaludica subsp. nov. was given earlier based on 5 eggs examined using a scanning electron microscope ( Kolesnichenko & Kotlobay 2020). Additional material made it possible to more accurately identify real diagnostic features.
The egg is the largest among the representatives of the lutko group. It is oval and noticeably elongated in the dorso-ventral direction ( Kolesnichenko & Kotlobay 2020). The height of the egg varies greatly from 761.0 µm to 855.0 µm, the width is from 627.5 µm to 703.0 µm ( Table 3 View Table 3 ). The sculpture of the micropilar area is formed from four to five rows of penta-hexagonal cells of various lengths and widths. The diameter of the micropilar rosette in the widest part varies from 78.0 µm to 90.0 µm. The micropilar rosette is formed by 7–9 (in rare cases 10) primary quadro-pentahedral cells with a width from 6.0 µm to 15.0 µm and a length from 9.0 µm to 27.5 µm. The micropile is rounded in shape, with an average diameter of about 8.0 µm. Lateral longitudinal ribs from 26 to 30 limit the micropile area and fall to ⅓ of the egg surface. The transverse ribs are expressed in the first third of the egg surface. Below the lateral ribs, the chorion is relatively smooth.
In general, the eggs of M. timandra binaludica subsp. nov. are similar to those of the nominate subspecies. A distinctive feature of the morphology of the eggs of the new subspecies is their well-pronounced elongated-oval shape. In the nominate subspecies, the egg shape is also oval, but not so elongated dorsoventrally. In addition, the eggs of M. timandra binaludica subsp. nov. are the largest, their maximum height is 850.0 µm, while the height of those of the nominate subspecies is about 700.0 µm.
Biology
The biology of the new subspecies in the Kuh-e-Binalud mountains and the early stages were described in detail by the authors earlier ( Kolesnichenko & Kotlobay 2020).
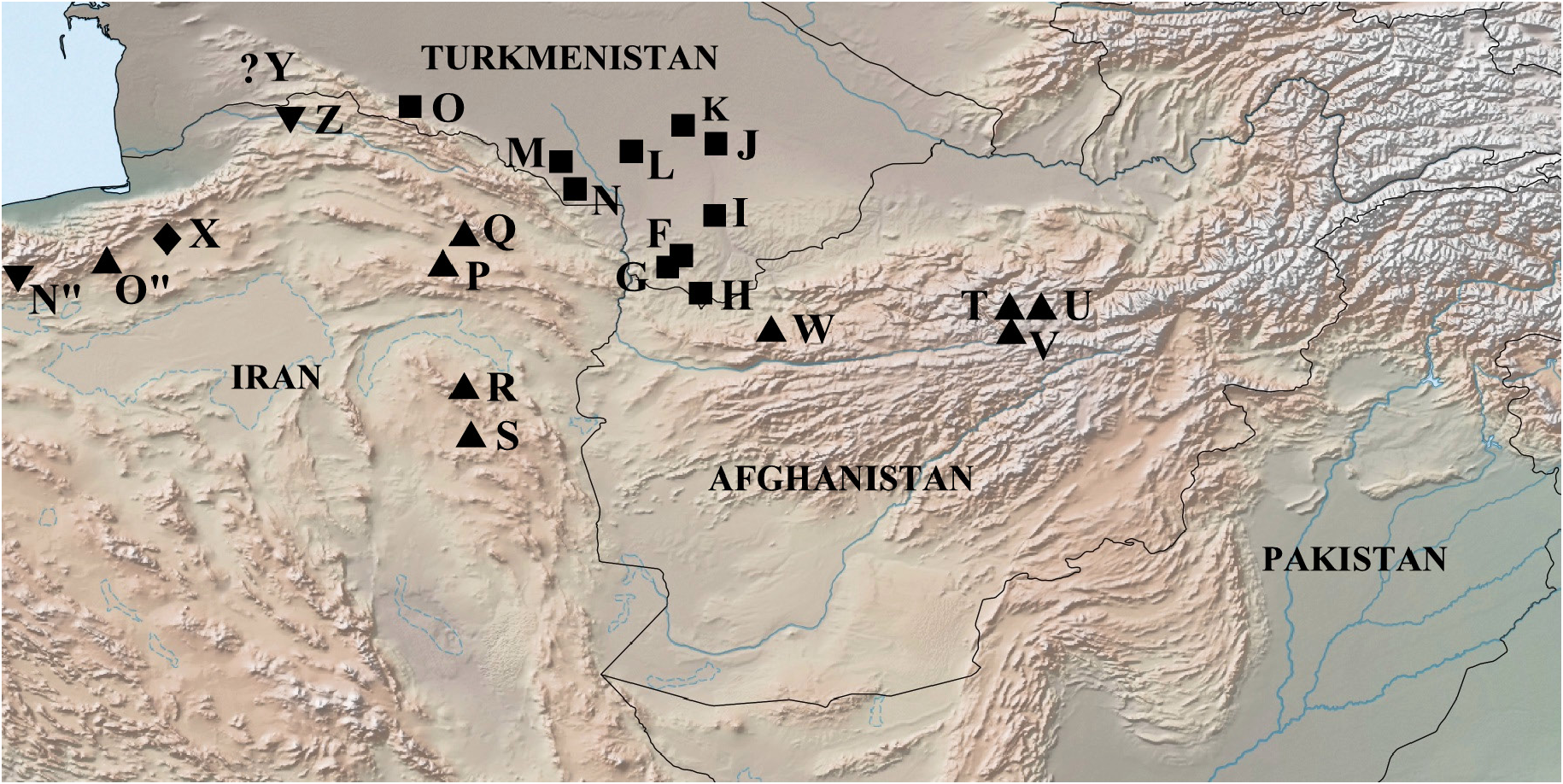
Distribution ( Fig. 14 View Fig )
Iran: Turkmen-Khorassan Mts, Kuh-e-Binalud Ridge, Kayen Mts; Afghanistan: Band-e-Amir.
The described subspecies M. timandra binaludica subsp. nov. undoubtedly should also include specimens of M. timandra ( M. lutko mimetica from the authors), caught by V. Eckweiler in Iran on Kuh-e-Binalud Ridge in the vicinity of Zoshk and Firizi ( Tshikolovets et al. 2014). The butterflies on the published photos ( Tshikolovets et al. 2014: tables lxi, 1–2) have an external similarity with the specimens of M. timandra binaludica available to us from the same place, caught at the same time by the same collector. The butterflies from the Bandi Amir Valley, identified by the authors ( Tshikolovets et al. 2018) as M. mimetica , should also be attributed to the described subspecies M. timandra binaludica .






| SDM |
Stroud and District Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Nymphalinae |
|
Genus |