
Gaucha piranauru, Souza & Ferreira & Carvalho, 2021
|
publication ID |
https://doi.org/10.11646/zootaxa.5061.3.9 |
|
publication LSID |
lsid:zoobank.org:pub:F3096BAE-FF30-4351-B488-DA98BF595F27 |
|
DOI |
https://doi.org/10.5281/zenodo.5700159 |
|
persistent identifier |
https://treatment.plazi.org/id/E723040D-5E16-D45C-9CC5-7C44FC53FD94 |
|
treatment provided by |
Plazi |
|
scientific name |
Gaucha piranauru |
| status |
sp. nov. |
Gaucha piranauru , new species
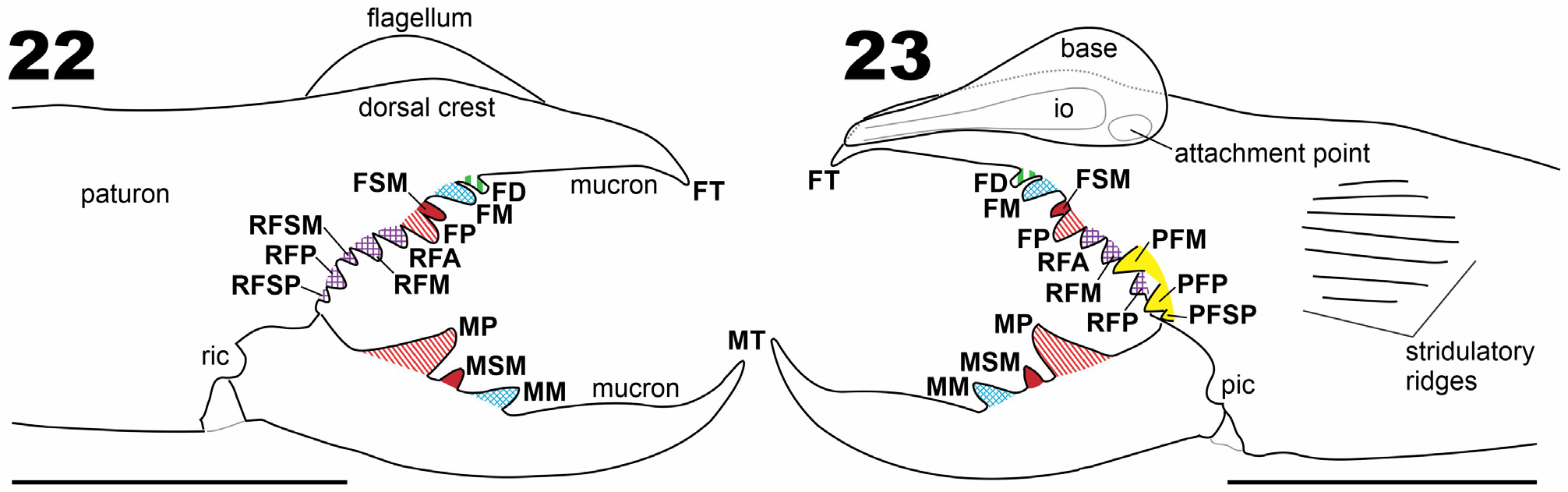
Figs 1–23 View FIGURE 1 View FIGURES 2–7 View FIGURES 8–13 View FIGURES 14–17 View FIGURES 18–21 View FIGURES 22–23 ; Tab. 1 View TABLE 1
Type material. Holotype: male from BRAZIL: Minas Gerais: Rio Pardo de Minas, Vale do Rio Peixe Bravo , 16°04’41.36”S, 42°43’42.53”W, 09.iv.2016, R. L. Ferreira leg., pitfall trap (ISLA 15684) GoogleMaps . Paratypes: 2 ♂ ( CHNUF-PI 4063 and ISLA 15683), same data of holotype, and GoogleMaps 1 ♀ ( ISLA 15685; Fig. 1 View FIGURE 1 ), same locality and collector of the holotype, 22.ii.2015, hand collection GoogleMaps .
Etymology. The epithet piranauru is formed by a combination of words: “ pira ” which means “fish” and “ naurú ” which means “angry”, both words coming from the Tupi-Guarani (Brazilian Indian languages). Therefore, piranauru means ‘‘angry fish”—in Portuguese, “peixe bravo”—in allusion to the name of the region where the species occurs (vale do rio Peixe Bravo). It is to be treated as a noun in apposition.
Diagnosis. Gaucha piranauru sp. nov. is a representative of the ibirapemussu species group, based on the longer mucron of the movable fingers of males (compared to females; Figs 8–13 View FIGURES 8–13 ), bearing a prominent and convex gnathal edge carina; by the absence of a FSD tooth ( Figs 22–23 View FIGURES 22–23 ); and by the long and slender cheliceral fixed finger mucron in males (compared to males of the fasciata species group). It can be distinguished from the remaining representatives of the ibirapemussu species group by the combination of the following characters: (1) reduced cheliceral fixed finger FD tooth, closely positioned and smaller than FM tooth; (2) cheliceral fixed finger FP tooth, larger than FM tooth; (3) cheliceral fixed finger with PFSP and RFSM teeth present; (4) cheliceral movable finger MM tooth well developed, larger than MSM tooth and smaller than MP tooth ( Figs 22–23 View FIGURES 22–23 ).
Description. Male holotype ( ISLA 15684). Color on 70% ethanol-preserved specimens: Propeltidium base color yellow, with yellowish-brown areas without well-defined borders in the central region and close to the edges (darker areas on anterolateral portion, close to the lateral lobes; Figs 2, 4 View FIGURES 2–7 ). Ocular tubercle brownish, darker around the eyes. Chelicerae with manus predominantly yellow with yellowish-brown areas dorsally and retrolaterally; fingers and teeth yellowish with reddish tips ( Figs 8, 10, 12 View FIGURES 8–13 ). Meso-, metapeltidium, and dorsal surface of opisthosoma with a three-dark-band design typical of the family ( Figs 2, 4 View FIGURES 2–7 ): tergites with median, longitudinal brown band, and paired, thinner lateral whitish bands, the latter with some brown pigment interspersed; pleural membranes with subdorsal brown and sub-ventral white bands; the white band of the opisthosomal pleural membrane with brown marks surrounding the socket of most setae, and brown pigment along the inter-segmental transversal vertices especially on posterior half ( Fig. 6 View FIGURES 2–7 ); sternites base color yellowish with some whitish pigment interspersed and lateral margins conspicuously darkened especially on two/three posteriormost sternites ( Fig. 7 View FIGURES 2–7 ), and preanal sternite entirely dark; anal plate dark except for dorsal whitish regions. Ventral surface of prosoma uniformly yellowish; sternum lighter than coxae. Pedipalps yellowish brown, with telotarsus darker than the rest of pedipalp ( Fig. 2 View FIGURES 2–7 ). Leg I brownish. Legs II to IV with proximal articles brownish on dorsal surfaces, lighter on ventral surfaces; tibia, basitarsus, and telotarsus darker and with more uniform coloration. Malleoli predominantly whitish with distal margin darkened. Morphology: Opisthosoma with ctenidia present, at least on 2nd to 4th post-genital sternites (spiracular sternite II and post-spiracular sternites I–II), a small number seen in spiracular II but more abundant in the others (see black arrows in Fig. 7 View FIGURES 2–7 ); ctenidia filiform and setiform, similar in thickness to the bifid setae (in male and female), but located on darker and larger sockets; ctenidia not seen in spiracular sternite I. A row of rigid hairs is present at 4 th postgenital sternite (white arrow in Fig. 7 View FIGURES 2–7 ). Pedipalp with all segments coated with bifurcated setae of different sizes; femur, basitarsus, and especially tibia with ventral set of very long setae, some of them as long as the tibia; clubbed setae not seen under light microscopy; spiniform setae absent. Leg I: thin, without claws and spines. Walking legs: covered with abundant small- to medium-sized bifurcated setae, and a few longer setae. Legs II and III: tibia with pro- and retroventral rows of three spiniform setae each, retrolateral ones thinner than prolateral, and distal-most pair longer and more robust than others; basitarsus with row of three retroventral, and one distal subventral spiniform setae, in a 2.2.3 rather staggered pattern; telotarsus bi-segmented with pro- and retroventral rows of five and four spiniform setae respectively, in a 1.2.2/2.2 pattern. Leg IV: tibia with row of five spiniform setae on proventral surface and single distal spiniform seta on retroventral surface, distal pair longer and slightly more robust; basitarsus with row of five proventral and one distal retroventral spiniform setae, in a 1.1.1.1,2 pattern; telotarsus bi-segmented with incomplete (ventral) segmentation on first (basal) tarsomere, with pro- and retroventral rows of six spiniform setae each, in a 2.2.2-2/2.2 pattern. Chelicera ( Figs 8–11 View FIGURES 8–13 , 14 View FIGURES 14–17 , 22, 23 View FIGURES 22–23 ): Fixed finger with all three primary teeth (FP, FM, and FD) and one secondary FSM tooth; FM columnar; FD reduced in size and with square apex and positioned very close to FM; FP longer than other teeth in the finger; FM larger and higher than FD, such that FP> FM >> FD; FM and FSM separated by a diastema. Fixed finger FSD secondary tooth absent. Fixed finger retrolateral fondal series composed of five teeth (RFA ≈ RFM> RFSM <RFP >> RFSP) and prolateral fondal series composed of three teeth (PFM ≈ PFP >> PFSP); PFM and PFP separated by a diastema. Fixed finger prodorsal carina straight along most of its length; mucron remarkably long and predominantly straight, not markedly thin, and with the apex (FT tooth) slightly curved towards the venter. Movable finger MP tooth pronounced, markedly higher than MSM and MM; MM larger than MSM, both teeth slightly higher than broader. Chelicera, prolateral surface with carpet-like field of barbed and bristle-like promedial ( pm) setae covering the distalmost third of manus ( Figs 8, 10 View FIGURES 8–13 , 15 View FIGURES 14–17 ). Flagellum drop-like, moderately inflated posteriorly and medially, with distal part narrow and tubular in appearance ( Figs 15, 16 View FIGURES 14–17 , 18 View FIGURES 18–21 ); ventral margin slightly sinuous ( Fig. 16 View FIGURES 14–17 ). Flagellum with conspicuous spicules along prodorsal margin ( Figs 18–20 View FIGURES 18–21 ), restricted to an area with a rugose tegument; both the rugose tegument and the spicules not extending to the retrolateral or to the prolateral surface ( Fig. 21 View FIGURES 18–21 ); apex of the flagellum reaching four-fifths of the mucron length from the base ( Figs 15–18 View FIGURES 14–17 View FIGURES 18–21 ), surpassing the mucron apex upon examination angle ( Figs 8, 10 View FIGURES 8–13 , 23 View FIGURES 22–23 ); apex narrow and with distinct spicules that share the rugose tegument with the flagellar base ( Fig. 17 View FIGURES 14–17 ).
Female paratype ( ISLA 15685; Fig. 1 View FIGURE 1 ). Similar to males with regard to coloration (except that female legs are brownish, darker at the tibia, basitarsus, and telotarsus; Figs 3, 5 View FIGURES 2–7 ) and most morphological aspects, but larger in size (morphometric values in Table 1 View TABLE 1 ). Chelicera on lateral aspect, fixed finger highest elevation at level of FD tooth ( Figs 12, 13 View FIGURES 8–13 ). Fixed finger robust, markedly curved towards the ventral side. Movable finger MP tooth slightly higher than MM tooth; MM higher than MSM ( Figs 12, 13 View FIGURES 8–13 ); movable finger retrolateral carina similarly developed to that of male .
Variation. All the chelicerae of males (n = 6 chelicerae) and female (n = 2) were confirmed to lack the fixed finger FSD tooth. Fixed finger primary teeth FM and FD are broken in the right chelicera (used for SEM; Figs 14–21 View FIGURES 14–17 View FIGURES 18–21 ) of one male paratype (CHNUFPI 4063). Fixed finger secondary tooth FSM is absent or broken in both chelicerae of the other male paratype ( ISLA 15683). Measurements of all males in Table 1 View TABLE 1 .
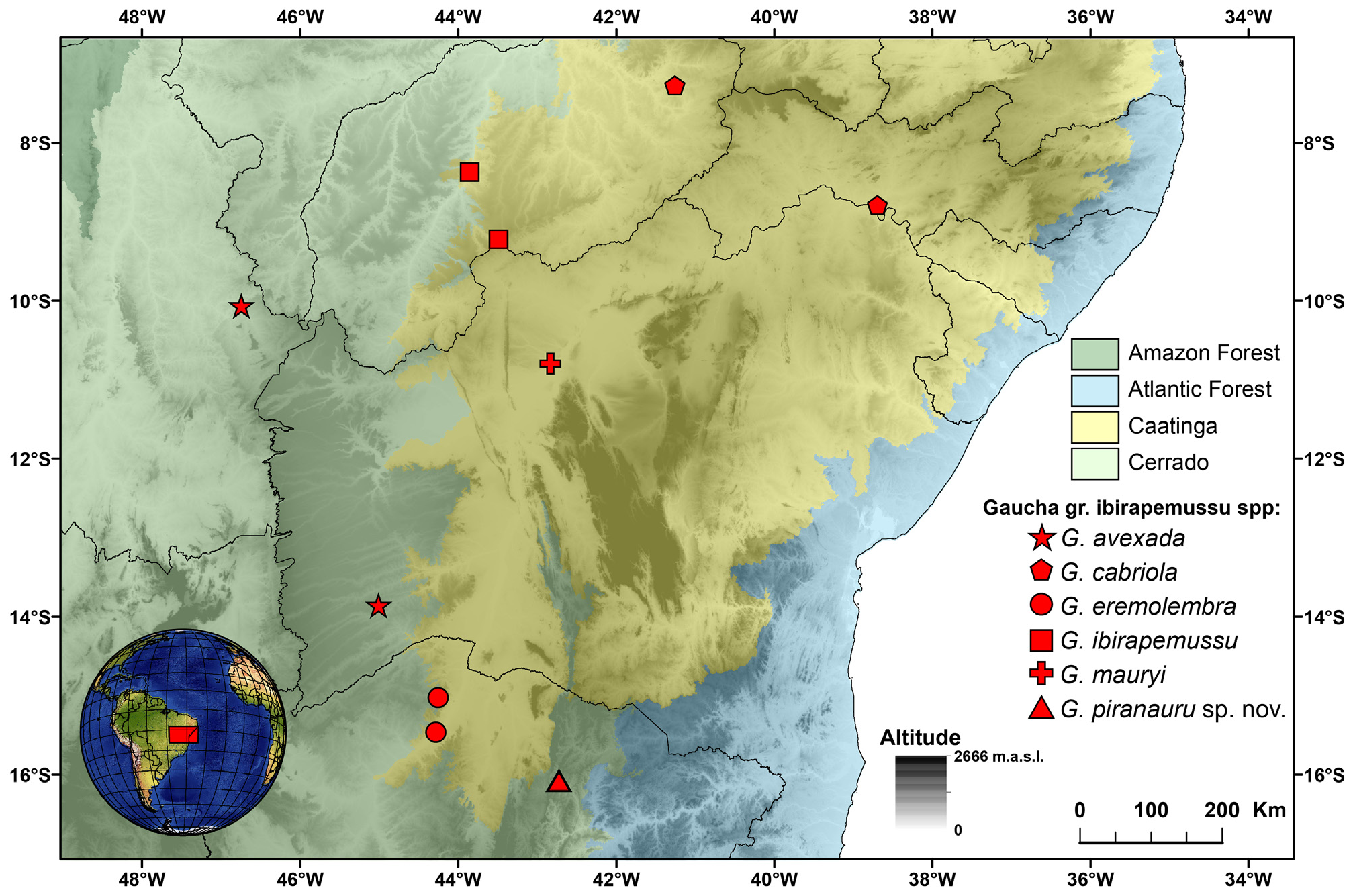
Distribution. Gaucha piranauru sp. nov. is known only from Rio Pardo de Minas, State of Minas Gerais, Southeastern Brazil ( Fig. 24 View FIGURE 24 ).
Natural history. The first specimen (female) was found during an expedition to the Vale do Rio Peixe Bravo region aiming at surveying cave fauna. In November 2015, at the end of the afternoon, a female was found while crossing a dirt road. In April 2016, a set of pitfalls was installed in the same area that the female was collected. The pitfalls were arranged in 100 meters linear transects and were installed 20 meters from each other (5 pitfalls per transect). Four transects were installed and the distance between them was 50 meters (hence, 20 pitfalls were installed, in an area of 20,000m 2). The pitfalls were filled with a saturated NaCl solution and were left in the field for three days. Three males were collected in pitfalls. Since only a single collection was conducted with pitfalls, it is not possible to currently address any aspect of the species’ autecology or behavior. However, the fact that the female was observed at the end of the afternoon may eventually indicate a diurnal/crepuscular activity, tough this merit further research. The area comprises a well-preserved Cerrado (Brazilian savanna) patch, in the northern state of Minas Gerais, Southeastern Brazil ( Figs 25–28 View FIGURES 25–28 ). However, it is important to mention that several areas have been deforested in the region, and the pristine Cerrado vegetation has been replaced by Eucalyptus crops, whose wood is used in industries. This ongoing deforestation process, allied to the potential risk of future mining activities (the region is rich in iron ore) raises concerns about the future of the area.






| R |
Departamento de Geologia, Universidad de Chile |
| MP |
Mohonk Preserve, Inc. |
| MM |
University of Montpellier |
| MSM |
Marine Science Museum, Tokai Univ. |
| FM |
Department of Nature, Fujian Province Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |