
Neoxorides montanus Oehlke, 1966
|
publication ID |
https://doi.org/ 10.5852/ejt.2020.680 |
|
publication LSID |
lsid:zoobank.org:pub:A8502F37-B3B5-49FC-9CBE-BA993BD426AA |
|
DOI |
https://doi.org/10.5281/zenodo.4329271 |
|
persistent identifier |
https://treatment.plazi.org/id/E26F87CB-FFA3-FFA6-FDB7-FE89CE24FB44 |
|
treatment provided by |
Valdenar |
|
scientific name |
Neoxorides montanus Oehlke, 1966 |
| status |
|
Neoxorides montanus Oehlke, 1966 View in CoL
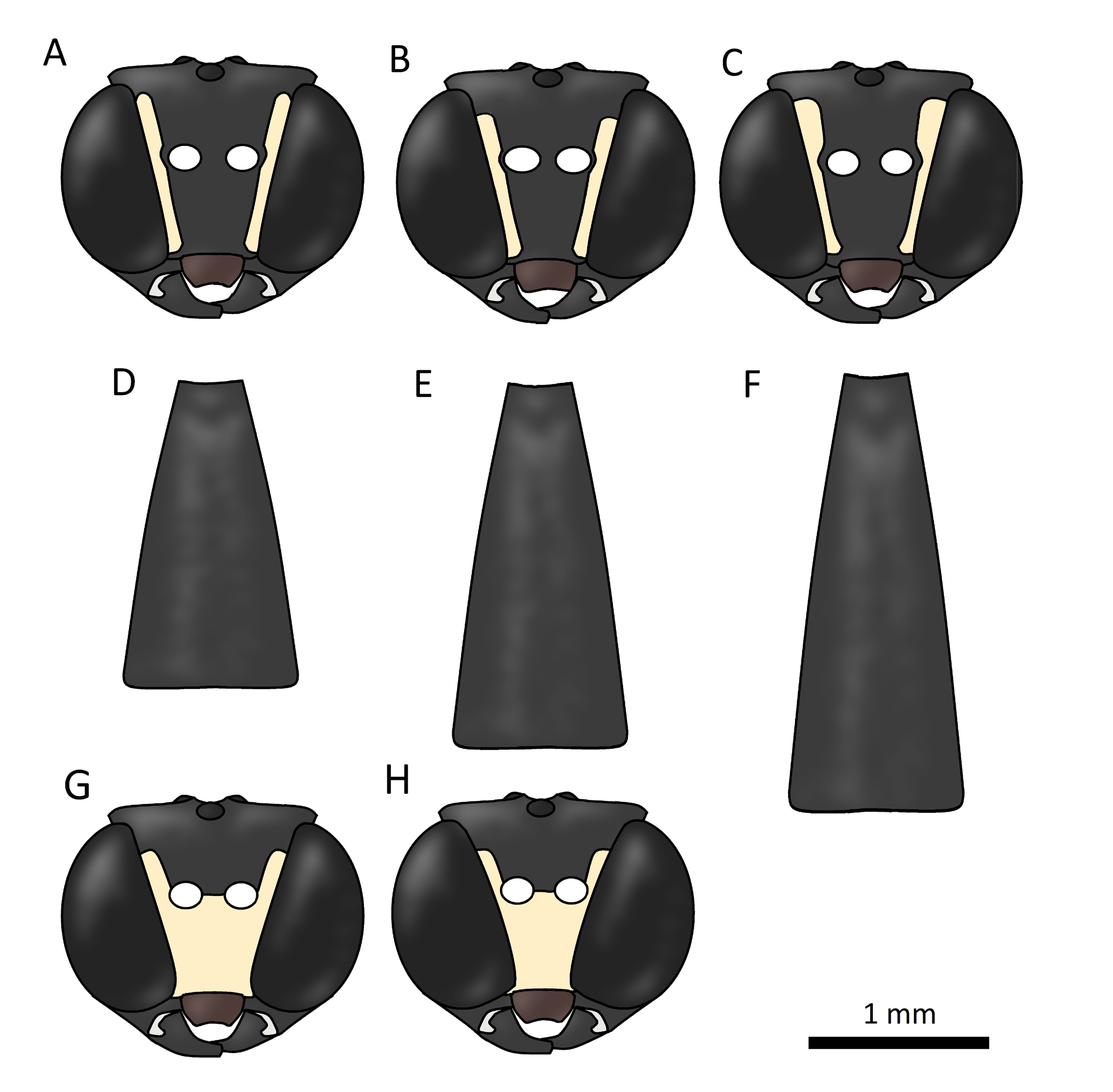
Figs 5G View Fig , 9C View Fig , F–G
Neoxorides montanus Oehlke, 1966: 889–890 View in CoL , fig. 3.
Neoxorides montanus View in CoL – Kasparyan 1981: 94.
Diagnosis
Neoxorides montanus is a species that is quite variable in size, with a fore wing length of 9–14 mm, on average slightly larger than N. collaris . The metasoma in the female is more elongate with the first tergite about 2.5 times as long as wide ( Fig. 9F View Fig ). The ovipositor in the female is usually as long as the metasoma and quite narrow and weak. In mounted specimens it is usually curved downwards, sinuate or curled up due to this weakness. The mid coxae in both sexes are largely yellow apically. The whitish spots along the inner orbits above the antennal scrobes are large and rounded in females ( Fig. 9C View Fig ) and the white lines on the sides of the pronotum are wide, usually as wide as the width of the fore femur ( Fig. 5G View Fig ). Neoxorides montanus is most similar to N. collaris and N. striatus sp. nov., but is distinguished by the characters given in the key to species and Table 2 View Table 2 .
Material examined
Holotype
CZECH REPUBLIC • ♂; ZMHB.
Paratypes
GERMANY • 3 ♀♀; ZMHB • 1 ♂; NHRS.
Other material
SWEDEN • 6 ♀♀, 2 ♂♂; MZLU, NJ, LW .
FINLAND • 4 ♀♀, 11 ♂♂; MZH .
FRANCE • 2 ♀♀, 2 ♂♂; NHMUK .
GERMANY • 3 ♀♀, 3 ♂♂; MR, NHMUK, MZH .
UKRAINE • 1 ♀; OV .
Ecology
Neoxorides montanus is connected to cerambycids feeding on conifers. It has been reared from larch along with the cerambycid Tetropium gabrieli Weise, 1905 (first author pers. obs.). Most records are from spruce Picea abies (L.) H.Karst. dominated forests, which possibly might indicate that the main hosts are Tetropium fuscum (Fabricius, 1787) and/or T. castaneum (Fabricius, 1787) ( Kenis & Hilszczański 2004) .
Distribution and status in Sweden
Most records are from the southern and central parts of the country, but it can probably be expected to occur also up in the north. Go, Hs, Sm.
Remarks
Among the distinguishing characters used to define and describe N. montanus , the relative length of the basal flagellomeres in the female and the shape of the male genitalia seem to be of limited use. According to the original description, the female of N. collaris is said to have the second flagellomere at most 5.2 times as long as wide while it is at least 5.5 times as long as wide in N. montanus . Given the natural variation in the shape of the flagellomeres (which are often oval in cross section) often amplified during preparation and preservation, this small difference is of no practical significance. The male of N. montanus usually has the parameres (and the hind margins of the tergites) more weakly sclerotized than in N. collaris and N. striatus sp. nov., which often makes them heavily deformed during storage or preparation. Newly hatched specimens or specimens of other species stored in ethanol also often display this deformation, making it hard to draw any definitive conclusions from the shape of the genitalia. We, therefore, recommend that the shape of the parameres is only used in combination with other characters. Apart from the shape of the male genitalia and the shape of the flagellomeres, the only diagnostic character used in the original description is the relative length of the metasomal segments. This seems to be a quite consistent character, but one should be aware that overlap frequently occurs with N. striatus sp. nov. and even N. collaris . During this study, it was also noted that the males of N. nitens and also N. striatus sp. nov. seem to be rather variable regarding the length of the metasomal segments ( Fig. 5 View Fig E–F). Both the holotype of N. montanus ( Oehlke 1966) and the lectotype of N. collaris ( Jałoszyński & Wanat 2014) are males, and this variation regarding the relative length of the metasoma could present a problem regarding the identity of the type specimens. However, all males of N. collaris studied by the first author have stouter metasoma, and all males of N. montanus have the more slender form, which indicates that these aberrant forms are rare. The additional colour characters of the lectotype of N. collaris and the holotype of N. montanus , respectively, i.e., the shape of the white spots along the inner orbits above the antennal scobes, also supports the validity of each species.
DNA barcode
The barcode of this species is separated from its closest barcode (in N. striatus sp. nov.) by 4.6% p-distance. The sequence is available on GenBank under accession number: MT 072691 View Materials (SK19_37).

| ZMHB |
Germany, Berlin, Museum fuer Naturkunde der Humboldt-Universitaet |
| MZLU |
Sweden, Lund, Lund University |
| NHRS |
Swedish Museum of Natural History, Entomology Collections |
| MZLU |
Lund University |
| LW |
Ivan Franko National University of Lviv |
| MZH |
Finnish Museum of Natural History |
| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Ichneumonoidea |
|
Family |
|
|
SubFamily |
Poemeniinae |
|
Genus |
Neoxorides montanus Oehlke, 1966
| Johansson, Niklas & Klopfstein, Seraina 2020 |
Neoxorides montanus
| Kasparyan D. R. 1981: 94 |
Neoxorides montanus
| Oehlke J. 1966: 890 |