
Limnonectes kong, Dehling, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4317.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:E82Ab46E-Cb3A-4513-808F-6C9Cf17Be372 |
|
DOI |
https://doi.org/10.5281/zenodo.6507987 |
|
persistent identifier |
https://treatment.plazi.org/id/DF4E87F0-FFEC-DE66-C3E1-72FCFCB0FD40 |
|
treatment provided by |
Plazi |
|
scientific name |
Limnonectes kong |
| status |
sp. nov. |
Limnonectes kong View in CoL sp. nov.
Rana conspicillata: Günther, 1872 (partim)
Rana kuhlii: Günther, 1874 (partim)
Limnonectes conspicillatus: Matsui et al. 2013 View in CoL (partim)
Holotype. NMBE 1059923, adult male, from Kubah National Park (1°36.411’N, 110°11.399’E; 290 m a.s.l.), Matang Range, Sarawak, Malaysia (Borneo); ColleCted 26 September 2009 by J. Maximilian Dehling and D. Matthias Dehling.
Paratypes. NMBE 1059917 (juvenile male), 1059918 (adult female), 1059919 (adult male), 1059920 (subadult male), all from Kubah National Park; NMBE 1059924, 1059926 (two adult males), 1059925 (subadult male), all from Bako National Park (1°42.684’N, 110°26.769’E), Sarawak, Malaysia; 1059927–1059929, 1059932 (four adult females), 1059930–1059931 (two adult males), all from Gunung Gading National Park (1°41.706’N, 109°50.455’E), Sarawak, Malaysia; ColleCted by J. M. Dehling and D. M. Dehling between 22 September and 18 OCtober 2009.
Referred specimens. BMNH 1947.2.29.24, adult female, from Matang Range, Sarawak, Malaysia (Borneo) (paraleCtotype of Rana conspicillata ); NMBE 1059921 (subadult female), 1059922 (adult female) from Santubong National Park; Sarawak, Malaysia, ColleCted by J. M. Dehling in OCtober 2009.
Molecular data. Holotype—NMBE 1059923 (GenBank ACCession # MF322505 View Materials ); Paratypes—NMBE 1059917 ( MF322509 View Materials ), 1059919 ( MF322506 View Materials ), 1059920 ( MF322510 View Materials ), 1059926 ( MF322507 View Materials ), 1059927 ( MF322508 View Materials ).
Diagnosis: Based on the presenCe of odontoids in the lower jaw (larger in males than in females), the enlarged, wide head, moderately depressed, stout body, short legs, thiCk and wrinkled skin, and the hidden tympanum, the new speCies is plaCed in the L. kuhlii group of the genus Limnonectes . It is distinguished from the other members of the group by the Combination of the following CharaCters: Large body size and marked sexual dimorphism with SVL 57.7–76.3 mm in adult males and 52.5–59.4 mm in adult females; head very large in males and markedly sexually dimorphiC, males (HW/SVL 0.41–0.48, HL/SVL 0.44–0.50) having wider and longer heads than females (HW/SVL 0.38–0.40; HL/SVL HL/SVL 0.39–0.43); head slightly longer than wide (HW/HL 0.93–1.00 in males, 0.92–0.97 in females); odontoids in males enlarged with a rounded knob-like tip; vomerine teeth in two highly elevated, large, broad posteromedially direCted ridges, separated from eaCh other by about half length of individual ridge; dorsolateral and transverse postorbital dermal folds absent; narrow, non-moveable dermal fringes between fingers along preaxial and postaxial sides of phalanges of hand; dorsal faCe of thigh and shank sparsely sCattered with enlarged, oval tuberCles; toes not fully webbed, leaving two phalanges on Toe IV and 1.5 phalanges on Toes II and III free.
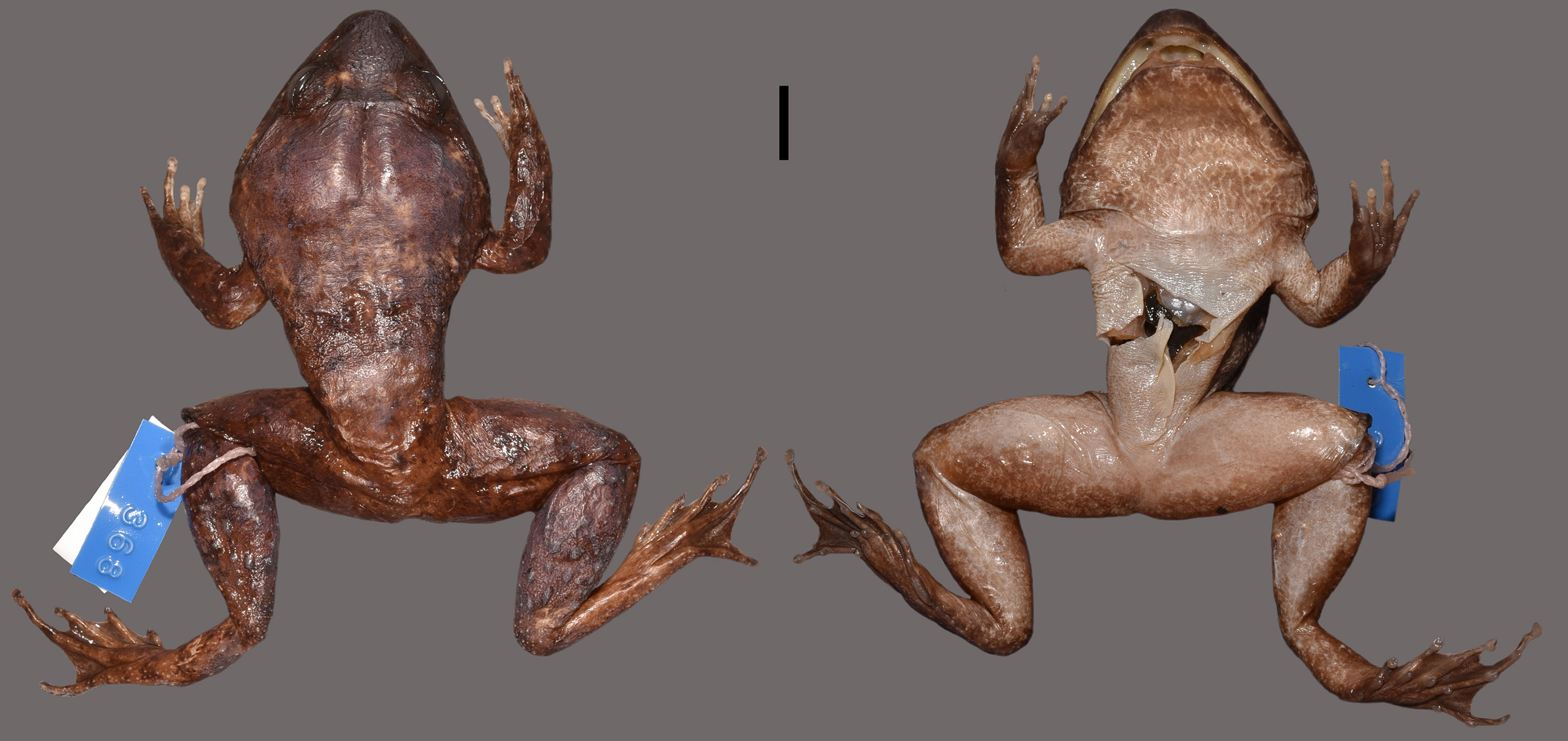
Description of the holotype. Adult male ( Fig. 5 View FIGURE 5 ); body large (SVL 68.3 mm; Table 2), very stout, widest posterior to head, strongly tapering to groin; head massively enlarged, very long and wide (HL/SUL 0.48, HW/ SUL 0.48), as long as wide (HW/HL 1.00); snout relatively short (SL/HL 0.34), aCuminate in dorsal view, rounded in profile, slightly projeCting beyond lower jaw, little wider than long (SL/EE 0.96); Canthus rostralis moderately expressed between eye and nostril, straight-lined; loreal region grooved; nostrils rounded, direCted dorsolaterally; situated Closer to tip of snout than to eye (EN/NS 1.20), separated from eaCh other by distanCe smaller than distanCe between eye and nostril (NN/EN 0.90); eyes direCted anterolaterally, protruding, small (ED/HL 0.24); eye diameter muCh shorter than snout length (ED/SL 0.71); interorbital distanCe muCh wider than upper eyelid (IO/EW 1.60) and wider than internarial distanCe (IO/NN 1.30); tympanum ConCealed under thiCk layer of skin, not disCernible; upper jaw with dentition; teeth on premaxilla muCh larger than those on maxilla, forming dense, transverse row; Choanae rounded; vomerine teeth in two highly elevated, large, broad posteromedially direCted ridges, separated from eaCh other by about half length of individual ridge ( Fig. 6 View FIGURE 6 ); mandibular symphysis thiCkened to large bony knob; odontoid proCess present Close to mandibular symphysis on both sides, direCted dorsally, large, long (OL/MH 2.1; OL/SVL 0.08) with blunt, rounded tip, fitting into deep rounded Cavities in upper jaw when mouth Closed ( Fig. 6 View FIGURE 6 ); tongue Comparatively short and broad, Covered with small pustules, most densely in anterior third.
Dorsal side of trunk, head, and limbs rugose; posterior half of trunk and dorsal faCe of thigh and shank sparsely sCattered with enlarged, oval tuberCles ( Figs. 4 View FIGURE 4 and 5 View FIGURE 5 ); supratympaniC fold ConspiCuous, Curved, running from posterior Corner of eye to anterior edge of fore limb insertion; ventral side with dense, weak, low wrinkles; transversal dermal fold on Chest between anterior edges of arm insertion.
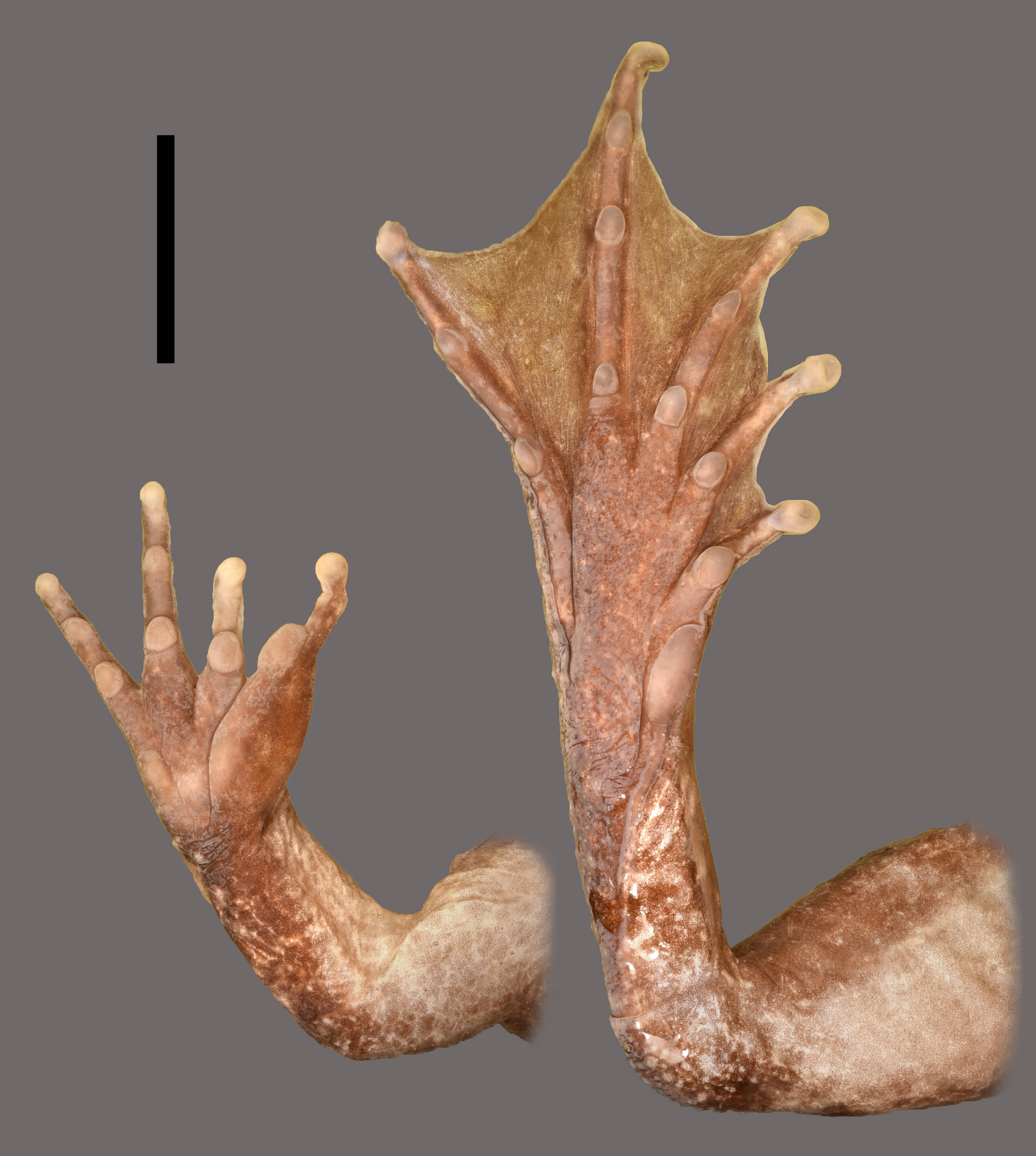
Forelimbs stout, short (ARM/SVL 0.42); hand relatively short (HND/SVL 0.23); tips of fingers rounded, swollen palmarly ( Fig. 7 View FIGURE 7 ); relative length of fingers: II <I <IV <III; subartiCular tuberCles rounded, well developed, numbering one on Fingers I and II, two on Fingers III and IV; tuberCles on Fingers I and II and proximal tuberCles on Fingers III and IV muCh larger than distal ones, highly elevated ( Fig. 7 View FIGURE 7 ); fingers without webbing but with narrow dermal fringes between fingers along phalanges from subartiCular tuberCles to disCs, most ConspiCuous on Fingers II and III, not moveable ( Fig. 7 View FIGURE 7 ); thenar tuberCle small, about two-fifth length and about one-sixth area of metaCarpal of Finger I in palmar view, oval, low; inner palmar tuberCle subequal in shape and size to thenar tuberCle, Covering proximal part of metaCarpals of Fingers II and III; outer palmar tuberCle somewhat narrower, but equal in length to thenar and inner palmar tuberCle, slightly more prominent, Covering proximal part of metaCarpal of Finger IV ( Fig. 7 View FIGURE 7 ); nuptial pad as large and distinCt glandular swelling on dorsal side of Finger I, with extremely minute asperities as regularly, widely sCattered roundish, flat glandular knobs on dorsal side of metaCarpal and preaxial side of proximal phalanx.
Hindlimbs sturdy, moderately long (LEG/SVL 1.42); tibiofibula Comparatively short (TFL/SVL 0.45), subequal in length to thigh (TFL/THL 0.97); heels not meeting eaCh other when knees flexed and thighs held perpendiCularly to median plane; distinCt, low transverse dermal fold at heel; prominent dermal fold along distal three-fourth of preaxial side of tarsus to proximal end of inner metatarsal tuberCle ( Fig. 7 View FIGURE 7 ); foot length about equal to tibiofibula length (FOT/TFL 1.01); relative length of toes: I <II <V <III <IV; tips of toes enlarged to small disCs, without CirCummarginal groove; subartiCular tuberCles well developed, rounded, highly elevated, numbering one on Toes I and II, two on Toes III and V, and three on Toe IV; proximal subartiCular tuberCles larger and more prominent than distal ones; pedal webbing formula I 1 /1.5 II 1 /1.5 III 1 /2 IV 2 /1+ V ( Fig. 7 View FIGURE 7 ); dermal flap on preaxial side of Toe I and postaxial side of Toe V from distal edges of metatarsal tuberCles to proximal ends of disCs ( Fig. 7 View FIGURE 7 ); inner metatarsal tuberCle very prominent, large and very elongate (length: 4.1 mm), about three-fourth length of Metatarsal I and slightly less than two-fifths length of Toe I; outer metatarsal tuberCle very small and low, rounded, hardly disCernible ( Fig. 7 View FIGURE 7 ).
Colouration in life. Dorsal side dark grey, loosely sCattered tuberCles with a light grey tip; flanks and sides of legs lightened to brownish light grey; ventral side greyish white; ventral side of head light grey with whitish retiCulated pattern ( Fig. 8 View FIGURE 8 ). Palmar side of metaCarpals and plantar side of metatarsals dark grey.
Colouration in preservative. Pattern and lightness of Colouration very similar to state in life but Colour has turned from grey to brown ( Fig. 5 View FIGURE 5 ).
Variation. The male paratypes are very similar in overall size (SVL 57.7–76.3 mm) and head proportion (HW/ SVL 0.41–0.47, HL/SVL 0.44–0.50) to the holotype. The females are smaller (SVL 52.5–59.4 mm) and have shorter and narrower heads (HW/SVL 0.38–0.40; HL/SVL HL/SVL 0.39–0.43) ( Fig. 9 View FIGURE 9 ). The odontoids are muCh larger in adult males (OL/SVL 0.053–0.087) than in females (OL/SVL 0.036–0.044). Extend of webbing does not differ between the sexes and is generally very Conservative without a marked deviation from the extent observed in the holotype. The tympanum is disCernible only in the smallest paratype (NMBE 1059917, juvenile male). The granulation on the surfaCe of the posterior trunk and the hindlimbs is most prominently developed in the largest males and in adult females ( Fig. 9 View FIGURE 9 ). The surfaCe of subadult males is muCh smoother. This suggests that these tuberCles are developed in fully mature speCimens only. Colouration varies between speCimens and some are light grey, other light brown dorsally in life, with darker grey or brown pattern Consisting of transverse interorbital bars, stripes on the limbs and along the supratympaniC fold ( Fig. 9 View FIGURE 9 ). Two paratypes have a ConspiCuous longitudinal yellowish line on the baCk along the spine ( Fig. 9 View FIGURE 9 b).
Morphological comparison. Based on the inComplete toe webbing, L. kong is easily distinguished from all desCribed Bornean speCies of the L. kuhlii group with fully-webbed toes, i.e. Limnonectes conspicillatus and L. mocquardi . Adult females of L. kong (SVL 52.5–59.4 mm) are smaller than females of L. conspicillatus (SVL 64.9–69.0 mm) and are usually larger than females of L. mocquardi (SVL 36.6–53.0 mm; Matsui et al. 2013; JMD, unpubl. data). Males of L. kong (SVL 57.7–76.3 mm) are usually larger than males of L. mocquardi (SVL 46.6– 58.2 mm; Matsui et al. 2013; JMD, unpubl. data). In addition, L. kong differs from L. conspicillatus and L. mocquardi in the tympanum being indisCernible (vs. ususally disCernible in L. conspicillatus and L. mocquardi ), the vomerine ridge being short and separated from eaCh other by half length of individual ridge (vs. longer and separated by less than width of individual ridge in L. conspicillatus ; shorter and separated by a little less than individual length in L. mocquardi ), the middle fingers having very narrow, non-moveable dermal fringes on the phalanges only (vs. somewhat broader fringes in L. mocquardi ; broad, moveable flaps present on phalanges and metaCarpals in L. conspicillatus ).
Of the speCies of the L. kuhlii group from Borneo with inComplete webbing, L. asperatus Inger, Boeadi & Taufik, 1996 (SVL of female 36 mm, adult males 34-38 mm), L. cintalubang Matsui, Nishikawa & Eto, 2014 (females 32–43 mm, male 45 mm), L. hikidai (females 33.8–40.4 mm, males 30.1–36.8 mm), and L. sinuatodorsalis Matsui, 2015 (females 34–38 mm, males 28–32 mm) are muCh smaller than L. kong (females 52.5– 59.4 mm, males 57.7–76.3 mm) and laCk the massive enlargement of the head in adult males ( Inger et al. 1996; Matsui & Nishikawa 2014; Matsui et al. 2014b; Dehling 2014; Matsui 2015). The new speCies differs in the extent of webbing from all fully webbed speCies from the Sunda region, i.e. Limnonectes kuhlii , L. selatan Matsui, Belabut & Ahmad, 2014 , L. sisikdagu MCLeod, Horner, Husted, Barley & Iskandar, 2011, and L. utara Matsui, Belabut & Ahmad, 2014 (MCLeod et al. 2011; Matsui et al. 2014a). Limnonectes deinodon Dehling, 2014 and L. khasianus have less extensive webbing on the toes (I 1.75/2+ II 1.5/ 3- III 1.75/3 IV 3 /1.5 V and I 1.25/2 II 1 +/2 III 1 +/ 2 IV 2 /1+ V, respeCtively, vs. I 1 /1.5 II 1 /1.5 III 1 /2 IV 2 /1+ V in L. kong ), are muCh smaller (SVL 34.2–46.8 mm and 46.5 mm, respeCtively, in males; 36.8–46.1 mm and 32.6–36.5 mm, respeCtively, in females, vs. 57.7–76.3 mm in males and 52.5–59.4 mm in females of L. kong ), and the odontoid proCesses of their males have a pointed tip (vs. blunt, rounded odontoid tip in males).
Molecular differentiation. The sequenCes we obtained from the type speCimens showed no or little variation between eaCh other (0.0–0.6% pairwise divergenCe). Comparison of the sequenCes with those deposited in GenBank Corroborated that L. kong is neither ConspeCifiC with L. kuhlii from Java, nor with any other desCribed speCies from the Sunda region for whiCh sequenCes are available ( Table 3). The interspeCifiC divergenCe between desCribed speCies of the L. kuhlii group, for whiCh moleCular data are available, is 6.5% or higher (Matsui et al. 2010, 2014a, 2014b). The sequenCe of L. kong was most similar to those of the sympatriC L. hikidai (7.7%) and “ L. asperatus ” (6.1%) from Bukit Raya National Park in Kalimantan. The identities of the speCimens from whiCh the sequenCes labelled “ L. asperatus ” stem need to be reassessed, as they show different levels of divergenCe from L. kong and from eaCh other. Some of them probably stem from misidentified speCimens. A sequenCe from Serasan Island labelled “ L. asperatus ” shows only 1.8% divergenCe from L. kong . This speCimen Could in faCt belong to L. kong , but this should be Confirmed through a morphologiCal re-examination..
The sequenCe of L. kong is identiCal with one of the 17 Bornean lineages reCently identified by Matsui et al. (2016), i.e. “lineage 8”. None of these 17 lineages has so far been attributed to either L. conspicillatus nor L. mocquardi . However, “lineage 6” of Matsui et al. (2016) from Matang, whiCh is identiCal with “’ L. 'kuhlii' Matang” of Matsui et al. (2010, 2014) and Matsui & Nishikawa (2014) Could be referable to L. conspicillatus ; and “’ L. 'kuhlii' Kinabalu” of Matsui et al. (2010) and Matsui & Nishikawa (2014) Could be referable to L. mocquardi . This needs to be Confirmed by morphologiCal examinations. SequenCes of these lineages differ by 7.1–7.7% and 9.3%, respeCtively from sequenCes of L. kong ( Table 3). Matsui et al. (2016) provided a phylogenetiC analysis of different lineages that are present on Borneo and their relationship to speCies from outside Borneo. SinCe L. kong does not represent an additional lineage, we refer to this resourCe for an analysis and depiCtion of the phylogenetiC relationships of L. kong .
Distribution. The type speCimens were ColleCted in Kubah, Gading, and Bako National Parks, and speCimens referred to L. kong based on morphologiCal CharaCters were ColleCted in Santubong National Park. All loCalities are in western Sarawak ( Fig. 10 View FIGURE 10 ). Matsui et al. (2016) listed sequenCes obtained from speCimens from Samunsam, Santu Bong, and Lanjak Entimau in western Sarawak, whiCh show little divergenCe (<1.0%) from the sequenCes of L. kong and are thus very likely ConspeCifiC. A sequenCe from Serasan National Park might indiCate that the speCies also oCCurs on the Natuna Islands, Indonesia. This island is loCated about 120 km and 160 km from Gading and Kubah National Parks, respeCtively, and less than 70 km off the Bornean Coast and was ConneCted to Borneo throughout most of the PleistoCene ( Voris 2000).
Ecology. The type speCimens were ColleCted from Calm seCtions of small streams (width 0.3–1.0 m) at elevations between 10 and 400 m a.s.l., where they sat in shallow water. Upon disseCtion, we found the male holotype to be heavily infested with many speCimens of an unidentified speCies of nematode whiCh were found enCapsuled on the surfaCe of the stomaCh ( Fig. 8 View FIGURE 8 ). StomaCh and intestines of the holotype Contained Chitinous remains of beetle head Capsules, body plates, and the hind legs of large loCusts.
Adult females ColleCted in September and OCtober Contained enlarged ova whiCh had a dark, almost blaCk animal pole and a Cream-Coloured vegetative pole. One paratype (NMBE 1059918) was found in a small Cavity at the stream edge ( Fig. 9 View FIGURE 9 a). The female repeatedly emitted a snarling, drawn-out, low-frequenCy Call that lasted between 0.5 and 3 seConds. A seCond speCimen Called simultaneously in the same way from farther away but we were unable to loCate it and determine whether it was a male or female speCimen.
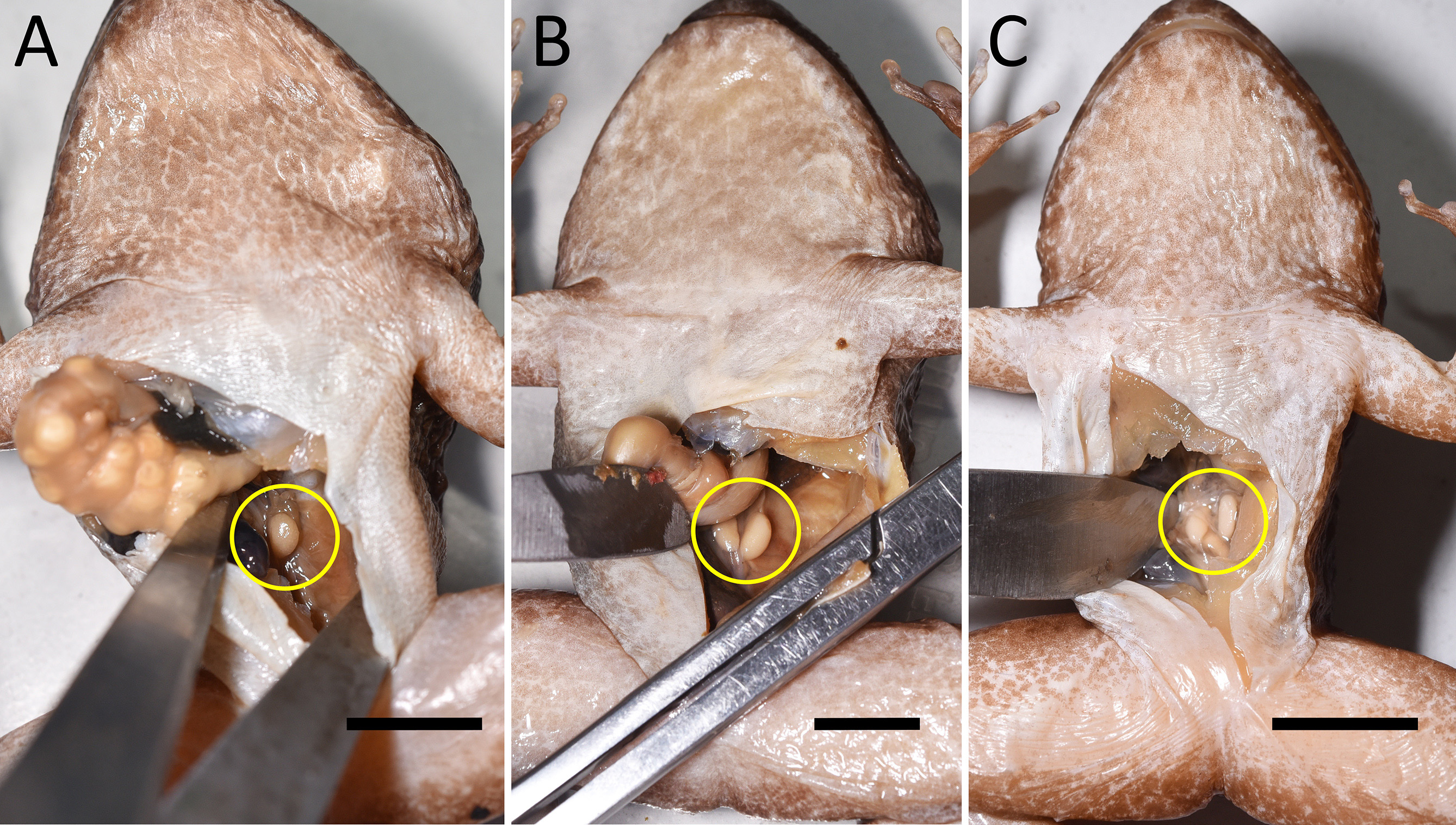
The testes of the largest male speCimens, inCluding the holotype, whiCh exhibit the most strongly developed seCondary sexual CharaCteristiCs (large head, large odontoid proCesses, tuberCles on posterior half of body), are relatively small (length x width 3.4 mm x 1.6 mm [holotype], 4.8 x 2.2 mm [paratype NMBE 1059924, Fig. 9 View FIGURE 9 ]; Fig. 11 View FIGURE 11 ). A smaller male (NMBE 1059925) that we determined to be “subadult” based on SVL, relative small head size, and small size of odontoid proCesses, has similarly sized testes (3.2 mm x 1.3 mm; Fig. 11 View FIGURE 11 ), and might in faCt already be mature.
Etymology. The speCies epithet is an allusion to the pronounCed sexual dimorphism exhibited by the new speCies. It resembles the male-biased dimorphism in great apes whiCh is most pronounCed in gorillas (Gorilla spp.). SinCe the new speCies is also among the largest of the L. kuhlii group on Borneo we name it after the fiCtional giant gorilla. The name is used as an invariable noun in apposition.





| NMBE |
Naturhistorisches Museum der Burgergemeinde Bern |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Limnonectes kong
| Dehling, D. Matthias 2017 |
Limnonectes conspicillatus:
| Matsui et al. 2013 |
Rana kuhlii: Günther, 1874
| : Gunther 1874 |
Rana conspicillata: Günther, 1872
| : Gunther 1872 |