
Odobenocetops, LEPTODON MUIZON, DOMNING & PARRISH, 1999
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2002.00015.x |
|
DOI |
https://doi.org/10.5281/zenodo.5700703 |
|
persistent identifier |
https://treatment.plazi.org/id/DE7C305B-FF9E-FA5A-F9F8-FDD3B023EF81 |
|
treatment provided by |
Carolina |
|
scientific name |
Odobenocetops |
| status |
|
ODOBENOCETOPS LEPTODON MUIZON, DOMNING & PARRISH, 1999
( FIGS 1–12 View Figure 1 View Figure 2 View Figure 3 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 View Figure 9 View Figure 10 View Figure 11 View Figure 12 , TABLES 1 AND 2)
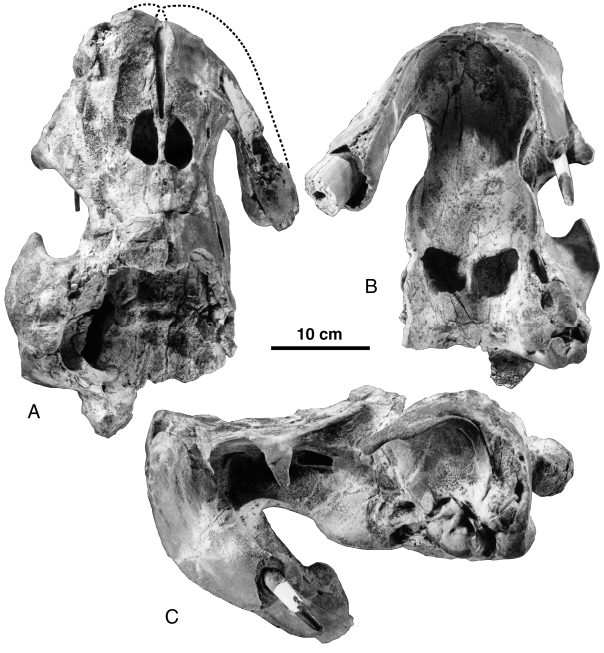
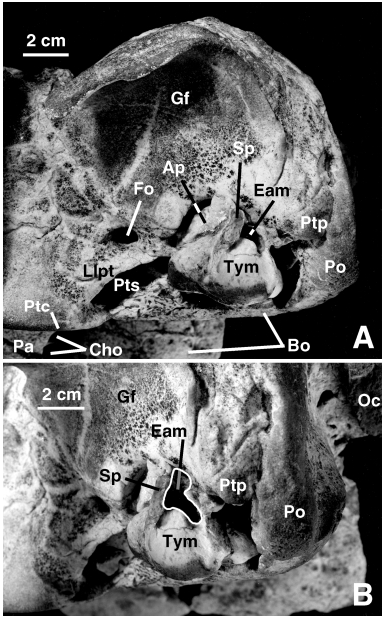
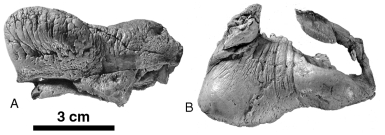
Holotype: SMNK PAL 2492 View Materials , an incomplete skull (lacking most of the dorsal and right sides of the braincase, right orbit, and right side of the right alveolar sheath), with the right ear bones and the atlas.
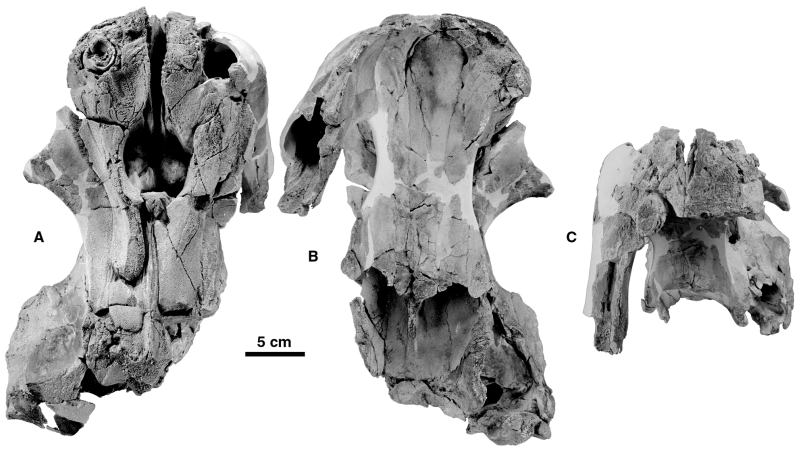
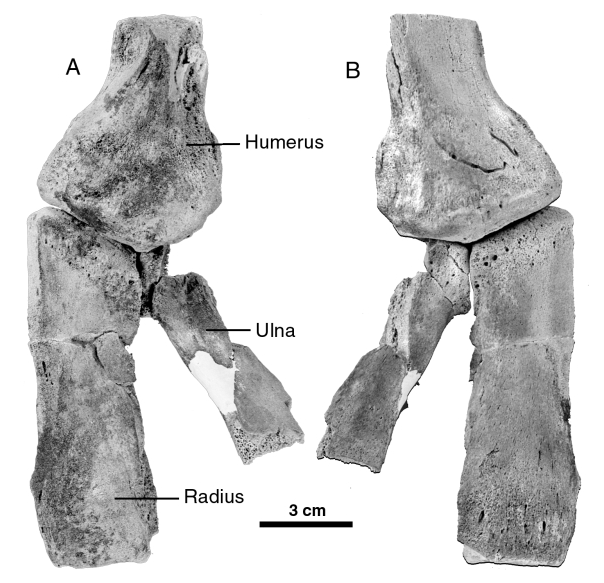
Referred specimen: MNHN SAO 202, partial skeleton with partial skull lacking the right tusk and with the alveolar sheaths very damaged. Most of the braincase has been weathered away but the left periotic and a partial tympanic are preserved in situ. The postcranial skeleton is very poorly preserved, comprising only a few hyoid fragments, 26 partial vertebrae, several rib fragments, the distal half of the left humerus, most of the left radius, a proximal portion of the ulna lacking the olecranon, and fragments of carpals and metacarpals. MNHN PPI 249, a right periotic is referred to O. cf. leptodon .
Etymology of the species name: from Greek: leptos (thin) and odon (tooth), in reference to the needle-like morphology of the large right tusk.
Diagnosis: Odobenocetops leptodon differs from O. peruvianus in the morphology of the snout, which is more rounded and wider in dorsal view; in the lack of large premaxillary foramina; in the presence of small depressions for premaxillary sacs in the premaxillae, anterior to the bony nares; in the presence of supplementary rostral bones at the anterodorsal apex of the snout; in the broader, deeper, and longer palate; in the anterior border of the palate, which is more U-shaped, while it is more V-shaped in O. peruvianus ; and in the straight (or slightly concave) anterodorsal border of the orbit, which is deeply concave in O. peruvianus .
Locality, Horizon and Age: the holotype and MNHN SAO 202 are from the SAO level of the Pisco Formation in Peru (at approximately km 540 of the Panamerican Highway south of Lima). The SAO horizon has been defined by de Muizon & Bellon (1980) but see also de Muizon (1981), de Muizon & DeVries (1985), and de Muizon & Bellon (1986). Its age spans approximately from 4 to 3 Ma; therefore, the specimens are early Pliocene in age. The SAO level of the Pisco Formation is slightly younger than the SAS level (5–4 Ma), which yielded the holotype of O. peruvianus and the third specimen referred to here as a female of O. peruvianus . MNHN PPI 249, an isolated periotic, is from beds of the Pisco Formation located at c. 3 km east of Yauca (these beds may be slightly younger than those of the SAO horizon and may belong to the late Pliocene).
Skull
Basicondylar length: 415e
Bizygomatic width: 150 ¥ 2 = 300e
Minimum width of the skull posterior to the postorbital processes: 70 ¥ 2 = 140e
Maximum width between the postorbital processes:
130 ¥ 2 = 260
Maximum width of both nares: 59e
Maximum length of the left naris: 53.5e
Width of the choanae: 104
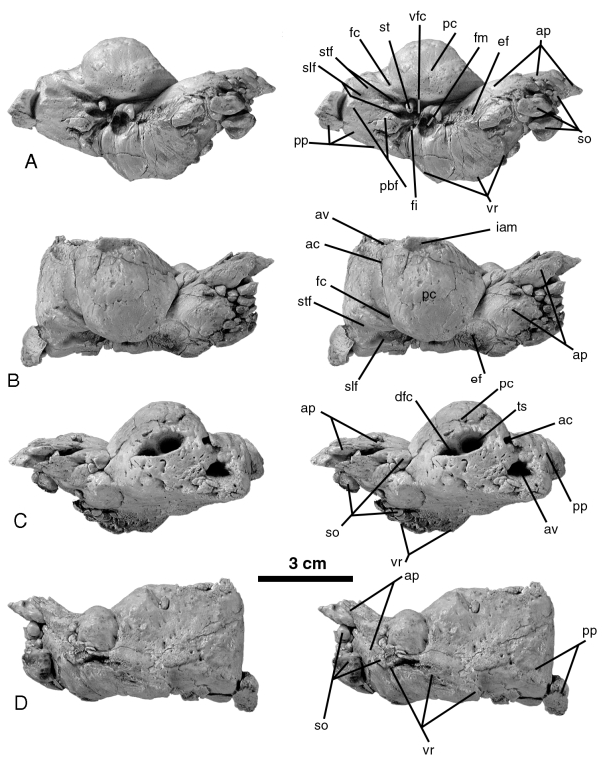
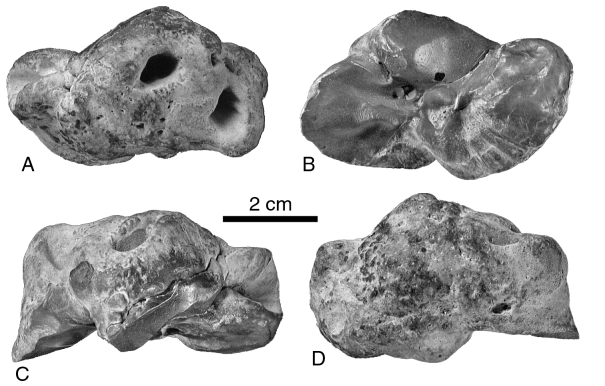
Periotic
Maximum width of the skull at the level of the exoccipitals: 123.5 ¥ 2 = 257e
Anteroposterior length: 77.5e
Maximum width (measured ventrally, approximately perpendicular to anteroposterior length: 42e
Maximum width between the pterygoid crests: 142 Width between the apices of the hamular processes: 112
Length of pars cochlearis from contact with anterior process anteriorly to medial edge of aqueductus cochleae (measured dorsally): 29.5
Tympanic
Length of the lateral lobe of the left tympanic: 54.2 Atlas
Maximum width of the pars cochlearis in medial view: 30.8
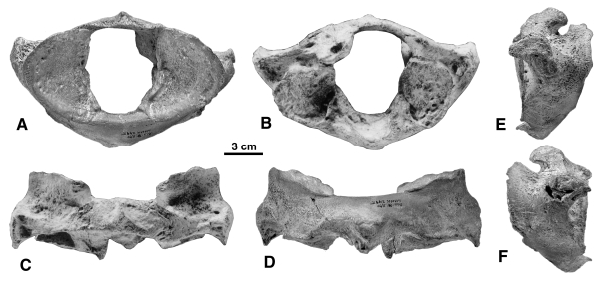
Maximum width of the atlas: 174
Length of internal auditory meatus: 14.3
Maximum height of the atlas at sagittal plane: 106 Maximum vertical diameter of neural canal: 64.5 Maximum horizontal diameter of neural canal: 56.5 Limb
Tympanic
Length of the involucrum (measured medially): 58.8 Malleus
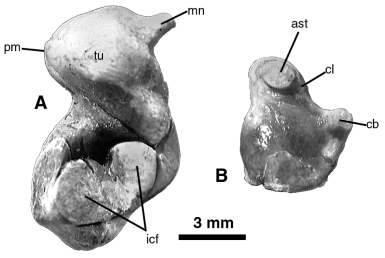
Maximum length in posteromedial view (cf. Fig. 10A View Figure 10 ): 11.4
Maximum length of the radius: 117
Anteroposterior length of distal end of humerus: 60.1 Mediolateral width of distal end of humerus: 43
Maximum width in posteromedial view (cf. Fig. 10A View Figure 10 ): 6.05
DESCRIPTION
The following description focuses on the differences between the two species of Odobenocetops . We refer to de Muizon et al. (2002) for the aspects of the cranial anatomy common to the two species.
GENERAL FEATURES
The general morphology of the skull of Odobenocetops leptodon is similar to that of O. peruvianus in its basic pattern ( Figs 1–4 View Figure 1 View Figure 2 View Figure 3 View Figure 4 ). However, in dorsal view, the former differs from the latter in the morphology of the snout, which is much wider and more blunt. In anterior view the snout was more massive. Furthermore, if a melon was actually present, as hypothesized below, it is likely that the head of O. leptodon was more rounded than that of O. peruvianus . The skull of O. leptodon has the same bilobate outline as is observed in O. peruvianus , which is unique in a Neogene odontocete ( de Muizon et al., 2002). This condition, where the frontals and maxillae do not overhang the temporal fossa, is found in primitive odontocetes such as Xenorophus , Agorophius and Archaeodelphis and in archaeocetes because of the lesser telescoping of the skull ( Kellogg, 1936; Fordyce, 1981). In Odobenocetops it is regarded as a reversal of this typically cetacean telescoping.
In general, the skull of O. leptodon is very heavily built and the bone is much denser, thicker, and more solid than in other delphinoids, being almost osteosclerotic in its posteroventrolateral angle.
PREMAXILLA
The premaxillae of O. leptodon are built on the same pattern as those of O. peruvianus ( de Muizon et al., 2002) . They both present a large alveolar process housing one tusk, the right tusk (of the presumed male, which is the only sex represented) being much larger than the left. Since the left alveolar process is not preserved on the holotype of O. peruvianus (the only specimen known when this species was first described) and because there are no other mammals which exhibit pronounced cranial asymmetry, it was hypothesized that the processes sheathing the tusks were both of the same size and that, consequently, the left tusk (which was judged to be shorter than the right sheath) was not erupted. The holotype of O. leptodon , however, indicates that the left sheath was clearly smaller than the right and that the left tusk was actually erupted. It is therefore probable that the condition of O. leptodon was also present in O. peruvianus . In the holotype of O. leptodon , the left sheath is transversely flattened and lies almost in a parasagittal plane (it diverges only slightly laterally). The right sheath is thicker because of the larger size of the tusk and diverges more markedly laterally. The region of the sheaths anterior to the tusks is strongly flattened and forms a posteriorly bent U-shaped lamina which borders the palate anteriorly. The anteroventral edge of this lamina is 0.5–1 cm thick, and presents longitudinal irregular ridges of bone to which was firmly attached the strong upper lip that was very probably present in O. leptodon as in O. peruvianus . The anterior and anterolateral side of each premaxillary sheath is a broad convex surface bearing several large grooves and foramina. These structures indicate an extensive vascularization of the anterior region of the skull, which is regarded here as related to the presence of a thick, muscular upper lip. In anterior view, in the sagittal plane, the snout is more elevated than in O. peruvianus and the surface of attachment of the upper lip is larger. Furthermore, the snout is wider in anterior view, and the anterior edge of the palate is grossly U-shaped rather than more Vshaped as in O. peruvianus . Therefore it is clear that the upper lip of O. leptodon was much larger than in O. peruvianus , indicating a larger vibrissal array and/or better suction ability in the former than in the latter.
Dorsally, the morphology of the premaxillae of O. leptodon presents the major characters which differentiate this species from O. peruvianus . These bones are much wider than in O. peruvianus , and the right premaxilla is more expanded laterally because of the greater size of the tusk ( Figs 2A View Figure 2 and 3A View Figure 3 ). There are no enlarged premaxillary foramina as are observed in the other odontocetes, including O. peruvianus . This absence is regarded as a loss in O. leptodon which, in this respect, is more derived than O. peruvianus . The premaxillary foramina transmit branches of the internal maxillary artery and maxillary nerve from the infraorbital foramen (internal to the orbit) to parts of the nasofacial muscles anteriorly and posteriorly on the rostrum (see e.g. Schenkkan, 1973: fig. 5, on Mesoplodon bidens ). In Odobenocetops these vessels and nerves are probably related to the irrigation and innervation of the strong upper lips and associated musculature as well as the possible vibrissae. In O. leptodon the canals conducting the nerves and vessels do not open dorsally but are clearly present. This is observed on the referred skull MNHN SAO 202 ( Fig. 3A View Figure 3 ), in which the apex of the snout is eroded above the pulp cavity of the tusks. The canals passed above the proximal extremities of the tusks and apparently (clearly observable on the right side) divided into several smaller canals perforating the premaxillae to reach the upper lip anteriorly. Ramifications of the premaxillary canals are also observable on the lateral side of the premaxilla at the level of the antorbital notch, where several large foramina are present. It is likely that irrigation and innervation for the pulp cavities of the tusks were also transported by the premaxillary canals.
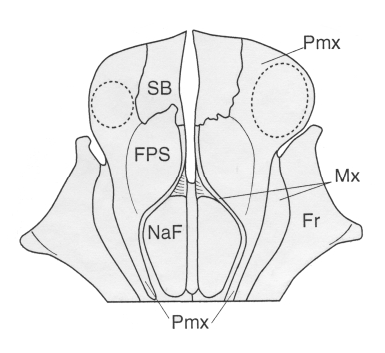
The premaxillae of O. leptodon bear a small concave area, anterior to the bony nares, where lay the premaxillary sacs of the air sac system ( Mead, 1975). These fossae for the premaxillary sacs are limited medially by the medial border of the premaxilla which overhangs the vomerian gutter; they are limited anteromedially by the suture with the rostral supplementary bone (see below); and anterolaterally they are medial to the posterior edge of each tusk alveolus ( Figs 2A View Figure 2 , 3A View Figure 3 and 5 View Figure 5 ). Fossae for premaxillary sacs are absent in O. peruvianus , where the space between the large premaxillary foramina and the anterior edge of the nares is so reduced that it was supposed that the premaxillary sacs were either very reduced or absent in this species ( de Muizon et al., 2002).
On the dorsal face of the skull the premaxillae of O. leptodon extend further posteriorly than in O. peruvianus . The posterior extremity of the left premaxilla is damaged on MNHN SAO 202, but it is clear that it lay slightly posterior to the posterior border of the nares. The posterior extremity of the right premaxilla lies about 5 cm posterior to the nares. On the holotype of O. peruvianus , the posterior extremity of the left premaxilla is located half-way between the anterior and posterior limits of the naris and on the right side (as well as on both sides of SMNK PAL 2491) it reaches the posterior edge of the naris.
The Pmx-Mx suture is external to the nares, the border of which is formed by the maxillae only, in contrast to the condition of O. peruvianus in which the Pmx-Mx suture is internal to the nares in the anterior two-thirds of their length. In their dorsal portions the premaxillae of O. leptodon are relatively flat and not as salient as is observed in O. peruvianus . This feature is clearly observed on the lateral edges of the nares and at the level of the antorbital notch, where the premaxillae markedly overhang the maxillae in O. peruvianus , whereas the relief of these regions is much smoother in O. leptodon .
The anterior sides of the premaxillae of O. leptodon (SMNK PAL 2492) bear strong foramina and grooves, which indicates an abundant vascularization related to the development of the upper lip. The premaxillae are much broader and much more elevated than in O. peruvianus , again indicating a larger and stronger upper lip ( Figs 1B View Figure 1 and 3C View Figure 3 ). The premaxillary sheaths, which house the tusks, are partly preserved on SMNK PAL 2492. The right is complete and the left is broken, apparently no more than 2 or 3 cm from the apex. Contrary to what was hypothesized in the case of O. peruvianus , the sheaths are not identical in size; the left one is clearly smaller and shorter than the right and reflects the size difference between the tusks. In lateral view, the right alveolar process forms an angle of c. 60° with the dorsal plane of the skull (from the vertex to the anterodorsal tip of the rostrum). The anteroventral edges of the sheaths (which include the anterior edge of the palate) are thickened and corrugated, as is observed in O. peruvianus , which indicates attachments for strong, possibly horny, connective tissue for the attachment of the upper lip. This narrow edge also bears (in both species) a series of foramina, which could have been related to a fringe of vibrissae. In the style of feeding we have hypothesized, these could have allowed the animal to detect when the front edges of the sheaths were in contact with the sea floor.
The anteroventral crest of the premaxillae ( Figs 1B View Figure 1 and 3C View Figure 3 ) has a very wide U-shape whereas it is more V-shaped in O. peruvianus . This morphology is the result of the widening of the snout and is possibly related to the presence of the supplementary bones described below. The U-shaped anterior border of the palate is not perpendicular to the sagittal plane. In other words, in anterior view, if the sagittal plane is positioned vertically (based on pterygoids, basioccipital and vomerian groove), the plane of the anterior crest of the palate dips towards the right side of the skull. This is because the anteroventral edge of the smaller and less laterally divergent left sheath is dorsal to (more elevated than) that of the right sheath. This condition implies that when the animal was feeding (i.e. with the anterior border of the palate parallel to the sea floor; see de Muizon, 1993b and de Muizon et al., 2002), the sagittal plane was inclined toward the left side of the animal (i.e. dipping toward the right side). Therefore, because of the asymmetry of the premaxillae, O. leptodon was feeding slightly inclined on left side.
The premaxillae form the anterior region of the palate as is observed in O. peruvianus . Although the palate of O. leptodon is much broader than in O. peruvianus , the area occupied by the premaxillae in this part of the skull is only slightly larger than in O. peruvianus (see below).
ROSTRAL BONES
One of the most striking differentiating characters of O. leptodon is the presence of two supplementary bones (one on each side) anterior to the premaxillae at the anterodorsal apex of the snout. These bones are apparently appressed against the medial edges of the premaxillae and lie partly on the dorsal part of the skull and partly on the anterior slope of the snout. The dorsal portion is narrower than long but broadens anteriorly. It is limited posteriorly by the fossa for the premaxillary sac of the premaxilla, while medially it has a small contact with the medial portion of the maxilla on the dorsal face of the rostrum ( Figs 1B View Figure 1 , 2A View Figure 2 , 3A View Figure 3 and 5 View Figure 5 ). Within the vomerian groove, the rostral supplementary bone is bordered by the medial portion of the maxilla, but the edge of the groove is formed by the rostral bone itself. The lateral suture is directed anteroposteriorly and is located just medial to the tusk alveoli. The anterior portion is not observable on MNHN SAO 202 but it is in relatively good condition on the left side of SMNK PAL 2492. It is also relatively narrow and forms a strip of bone overlying the premaxilla, tapering ventrally, and reaching (or almost reaching) the ventromedial border of the premaxilla at the anterior edge of the palate. Its medial suture is with the vomer. These supplementary bones are in fact wedged (as far as the dorsal portion is concerned) between the tusks laterally and the vomer and maxilla medially. They are partly responsible for the breadth of the snout in O. leptodon and tend to separate the proximal extremities of the tusks and alveoli (i.e. the premaxillae). As a probable consequence of their presence, the tusks are less divergent distally than in O. peruvianus and the anterior border of the palate is Ushaped in O. leptodon rather than more V-shaped as in O. peruvianus . Apparently, as is observed on the left premaxilla of SMNK PAL 2492, the rostral bones were anterodorsally protruding, forming an elevated prominence at the apex of the snout. This condition is not observable on MNHN SAO 202 since the bones have been partly eroded in their apical portions.
These supplementary bones could be ossa prenasalia, cartilage bones which appear late in development (mainly in postnatal life) and which are not uncommon in mammals. They are paired or unpaired bones which articulate with the nasals or premaxillae and which are always related to the presence of a rhinarium ( Rhynchocyon, Sus , Talpa, Chiroptera ) or a proboscis ( Starck, 1967). In the case of O. leptodon the supplementary bones present at the anterodorsal apex of the snout could be related to the thick and powerful upper lip used by Odobenocetops to forage on the bottom in search of food ( de Muizon, 1993a;b; de Muizon et al., 2002). They could also indicate of the presence of a very sensitive tactile organ, with or without vibrissae (see de Muizon, 1993a), much more developed in O. leptodon than in O. peruvianus . It is likely that strong muscles for the upper lip of O. leptodon were attaching on the (?)prenasalia. However, it is noteworthy that the prenasalia of other mammals are generally not tightly articulated with the premaxillae and/or the nasals, and not distinctly paired.
If these supplementary bones are not prenasalia, another alternative which could explain the condition of O. leptodon is the presence of septomaxillae. These bones are present in crossopterygians, lissamphibians, squamates, in all nonmammalian therapsids, and in all the nontherian mammals except multituberculates. The septomaxilla is absent in marsupials and placentals except in Xenarthra ( Wible, 1990; Zeller et al., 1993). However, we are reluctant to interpret the supplementary bones of O. leptodon as septomaxillae, since the study of the development of the nasal capsule of odontocetes ( Klima & Van Bree, 1985; Klima et al., 1986; Klima, 1987) does not show any indication that such bones are present at any developmental stage in this group.
A third alternative is to regard the supplementary bones of O. leptodon as neomorphic elements different from prenasalia and septomaxillae, possibly related to the muscle attachment of the lip, which was probably much stronger than in O. peruvianus .
TUSKS
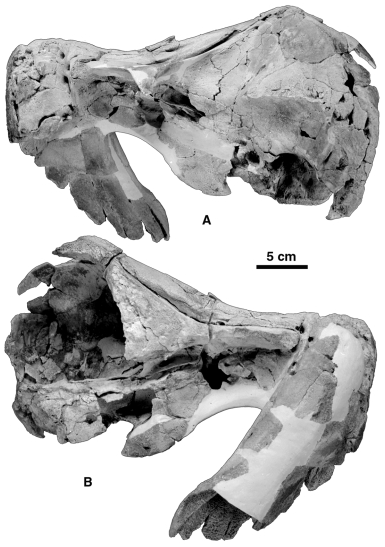
The tusks of O. leptodon are similar to those of O. peruvianus ( Figs 1A,B View Figure 1 ). They are much better preserved in SMNH PAL 2492 than in the holotype of O. peruvianus . The large right tusk is a straight, cylindrical, needle-like tooth. Its erupted portion (broken and worn during life) is 107 cm long and its alveolar portion (as preserved) is approximately 28 cm long. Before the loss of its original tip, it could easily have been 30 cm longer. Therefore, the right tusk of O. leptodon is much longer than what was estimated for O. peruvianus , in which we suggested that it was possibly no longer than 55 cm with a 15–20 cm long erupted portion. It is therefore probable that the long tusk of O. peruvianus was longer than previously suspected (but see below). In O. leptodon as in O. peruvianus , the section of the tusk is elliptical and the diameters diminish towards the apex. At the alveolar border the diameters are 50 mm and c. 36 mm, whereas at 8 cm from the broken and worn apex the diameters are 30 mm and 25 mm. The large tusk of O. leptodon is made of dentine only (it has no enamel) and bears longitudinal striae as in O. peruvianus . At the apex of the tusk is a long wear facet which faces ventrally. It is 8 cm long and convex ventrally. The facet is strongly oblique and its posterior edge does not present a marked limit with the ventral border of the tusk. The pulp cavity is wide open at the proximal extremity of the tooth, which indicates continuous growth. At the proximal extremity of the erupted portion (2 cm distal to the alveolar border) the diameters of the pulp cavity are 24 mm and 13 mm. Considering that the pulp cavity is almost as wide as the tooth itself at the proximal extremity of the tusk, and if (as in O. peruvianus ) the cavity was conical, the pulp cavity of O. leptodon should have been approximately 60 cm long, which is more than twice the length measured in O. peruvianus (23 cm). Hay & Mansfield (1989) stated that the pulp cavity of the narwhal tusk is conical at the root. However, ‘ 10–20 cm from the base it narrows to a canal 0.5 cm in diameter which passes nearly to the tip of the tusk’ ( Hay & Mansfield, 1989: 160). These authors, citing Porsild (1922), also stated that in very old males the pulp canal is nearly occluded. Therefore, if the age of the animal is unknown, the length of the pulp cavity is probably not a good criterion to estimate the length of the right tusk of O. peruvianus . The apex of the tusk of the holotype of O. leptodon shows an open cavity. Since the wear facet clearly indicates the apex of the tooth, this canal had to be separated from the pulp cavity. In fact, a radiograph of the tusk shows that 35 cm from the apex, this apical canal is probably closed by dentine. It is likely that when the tooth was unworn the pulp cavity extended almost to the apex, as is observed in the living narwhal.
The small tusk is not complete in either specimen of O. leptodon , but its apex is preserved on SMNK PAL 2492, although separated from the intra-alveolar portion of the tooth. Therefore, on the specimen ( Figs 2B,C View Figure 2 ) the reconstructed length of the tusk is an estimate, although probably relatively close to the actual length. The total length of the tusk was approximately 25 cm and the intra-alveolar portion approximately 20 cm. It is unlikely that more than 5 cm of the tusk were erupted. The eruption of the small tusk is corroborated by the presence, at the apex, of a small wear facet, which is grossly parallel to the anterior U-shaped border of the palate and therefore to the sea bottom in feeding position. The section of the small tusk is also oval and the diameters at its proximal extremity are 22 mm and 18 mm. At the extremity of the preserved portion of the alveolar part of the tusk (which is close to the alveolar border) the diameters are 18 mm and 14.5 mm. At the level of the proximal extremity of the wear facet (approximately 1 cm from the apex of the tusk), the diameters are 10 mm and 11 mm. The pulp cavity of the tusk is similar to that observed in O. peruvianus : it is small, conical, and approximately 1 cm deep, partially filled with a knot of irregular dentine on MNHN SAO 202. In the narwhal such a knot on the unerupted small tusk (right in the male, and both in the female) is produced when the growth of the tooth stops, generally prior to sexual maturity.
MAXILLA
The maxilla is reduced as in O. peruvianus , and is basically similar to that of this species. It does not participate in the construction of the bony palate, which has no teeth other than the tusks. In MNHN SAO 202, as is observed in O. peruvianus , the portions of the maxillae posterior to the nares are narrow strips of bone and the right is wider than the left. A slight asymmetry of the bones of the face (especially the maxillae) is characteristic of odontocetes, in which the right bones are more developed than the left.
FRONTALS
The frontals of O. leptodon also have the same basic pattern observed in O. peruvianus . The supraorbital process is a large triangular plate overhanging an anterolaterally orientated orbit. The dorsal border of the orbit differs from that of O. peruvianus in being only slightly concave instead of deeply notched. The condition of O. leptodon probably precluded (certainly reduced) the dorsal binocular vision that was hypothesized by de Muizon et al. (2002) for O. peruvianus .
PALATINES AND VOMER
On the palate the vomer of O. leptodon is more developed than in O. peruvianus , in which it is already much larger than in the other odontocetes. It extends further posteriorly on the palate than in O. peruvianus , and as a consequence the palate is wider in its median region and its lateral edges are not so deeply notched. In the posterior region of the palate of the holotype, the palatines are not in contact medially (as is observed in O. peruvianus ); as a result, the vomer is visible continuously in the middle of the palate from the anterior crest to the basisphenoid. However, on the referred specimen MNHN SAO 202 the palatines are almost in contact and the condition of the holotype may be an individual variation. The palate of O. leptodon is longer than in O. peruvianus . This condition is mainly due to the fact that the posterior edge of the palate (between the apices of the pterygoid hamuli) is convex posteriorly in the former while it is deeply concave in the latter. Consequently, the palate in O. leptodon is much larger than in O. peruvianus in both length and width.
PTERYGOIDS
The pterygoids have a large ventrolateral crest as in O. peruvianus , probably for attachment of the pars lateralis of the pterygoid muscle. However, the apex of the pterygoid hamulus is smaller and not as salient posteriorly as in O. peruvianus (see below). The apex of the pterygoid hamulus contacts the medial lamina of the pterygoid. The latter is greatly thickened and incorporated into the basioccipital crest as in O. peruvianus (see below, the description of SMNK PAL 2491, referred to O. peruvianus ). This contact of the pterygoid hamulus with the enlarged medial lamina of the pterygoid forms a large foramen through which passes the eustachian tube.
The pterygoid sinus is extremely reduced when compared to most other odontocetes. It is about one-tenth the size observed in Delphinapterus , and if a functional sinus was actually present it was obviously vestigial. In fact, the lateral side of the Odobenocetops skull between the apex of the tympanic and the pterygoid hamulus is considerably shortened.
A small lateral lamina of the pterygoid is present. However, contrary to the condition in other odontocetes it does not articulate with the squamosal but with the alisphenoid. It forms the anterior border of the foramen ovale on the lateral wall of the skull. This condition is only superficial since internally the foramen ovale is fully enclosed in the alisphenoid as in the other eutherians.
ALISPHENOID
As in O. peruvianus , the alisphenoid is a very thick bone, which contrasts with the condition observed in the other delphinoids. The foramen ovale is well separated from the cranial hiatus and not confluent with it as in the holotype of O. peruvianus , in which it is probably broken (see, below, the description of SMNK PAL 2491, a specimen referred to O. peruvianus , in which the alisphenoid region is much better preserved than in the holotype, USNM 488257). The foramen ovale is enormous (two to three times the size observed in Delphinapterus ), as is observed in O. peruvianus . The mandibular nerve, which exits the skull through the foramen ovale, has eight branches in the dog ( Miller et al., 1964): the pterygoid nerve, the buccal nerve, the deep temporal nerve, the masseteric nerve, the auriculotemporal nerve, the mylohyoid nerve, the mandibular alveolar nerve, and the lingual nerve. It is a mixed nerve, sensory and motor. Of course the condition in Odobenocetops was not necessarily similar to that in the dog. However, it is noteworthy that these branches totally or partially innervate the tongue, the masticatory and digastricus muscles, the hyoid muscles, the cheeks, and the lips. Therefore, the size of the foramen ovale and the inferred large size of the mandibular nerve are consistent with the large size of the tongue, the upper lip and the jaw muscles as well as with their function as hypothesized by de Muizon (1993a,b) and de Muizon et al. (1999; 2002).
SQUAMOSAL
The major characteristic of the squamosal of Odobenocetops is the highly modified glenoid cavity and zygomatic process, described by de Muizon et al. (2002), which forms a large, anteroposteriorly orientated gutter-like cavity ( Figs 1B View Figure 1 and 6 View Figure 6 ). This region, although following the same basic pattern in the two species, shows some differences in O. leptodon . In the latter, the glenoid cavity is approximately 15% shorter anteroposteriorly. Furthermore, in lateral view, the posterior border of the glenoid cavity lies 3–4 cm from the lambdoid crest of the exoccipital, whereas in O. peruvianus the posterior edge of the cavity is at the level of the crest. The lateral wall of the glenoid cavity (or gutter), although concave ventrolaterally, is almost horizontal and not strongly recurved ventrally as is observed in O. peruvianus . Furthermore, the zygomatic process of the squamosal is apparently located in a higher and more anterior position on the skull. This condition is not easy to observe on the available specimens of O. leptodon , since the roof of the braincase is almost totally lacking in SMNK PAL 2492 and the process is broken in MNHN SAO 202. It is clear, however, that the anterior notch of the glenoid cavity of O. leptodon is closer to the lateral ridge of the frontal (the posterior extension of the posterior edge of the supraorbital process) than in O. peruvianus . This separation measures c. 5.1 cm in SMNK PAL 2492 and c. 6.3 cm in MNHN SAO 202 ( O. leptodon ); c. 9.5 cm in USNM 488257 and c. 9.2 cm in SMNK PAL 2491 ( O. peruvianus ). The squamosalparietal suture is especially clear on MNHN SAO 202. It is similar to that described for O. peruvianus , having the same unusual zig-zag shape. This is likely to represent an adaptation to strong muscular tensions applied to the bones during the suction feeding of Odobenocetops .

| SMNK |
Staatliches Museum fuer Naturkunde Karlsruhe (State Museum of Natural History) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.