
Odobenocetops, PERUVIANUS MUIZON 1993
|
publication ID |
https://doi.org/ 10.1046/j.1096-3642.2002.00015.x |
|
DOI |
https://doi.org/10.5281/zenodo.5700705 |
|
persistent identifier |
https://treatment.plazi.org/id/DE7C305B-FF8C-FA46-F99B-FD3DB5B4E8A4 |
|
treatment provided by |
Carolina |
|
scientific name |
Odobenocetops |
| status |
|
ODOBENOCETOPS PERUVIANUS MUIZON 1993
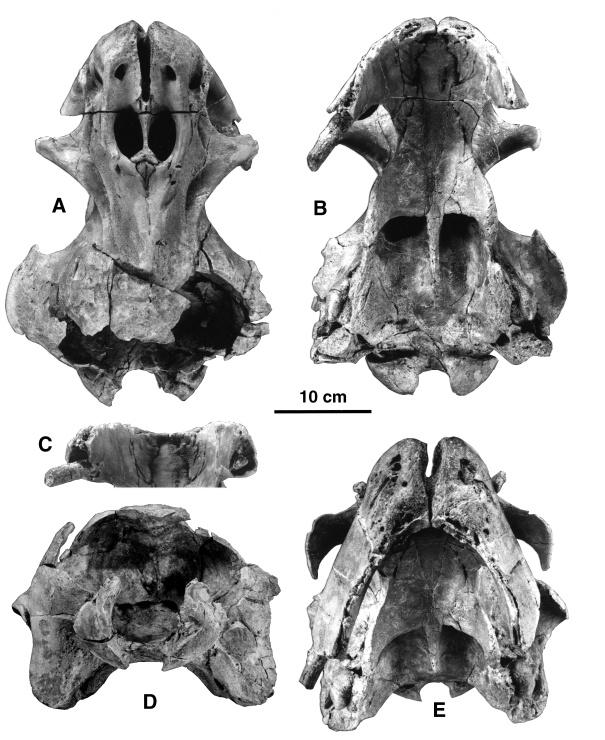
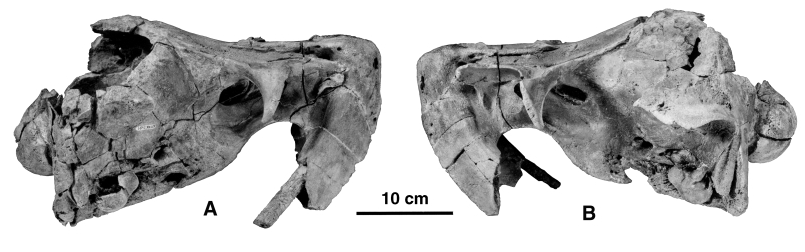
( FIGURES 13–15 View Figure 13 View Figure 14 View Figure 15 , TABLE 3)
A second skull of Odobenocetops peruvianus has been discovered in the earliest Pliocene beds of the Pisco Formation at Sud-Sacaco. This specimen is of special interest because it is more complete than the holotype and especially because it bears two small tusks similar in size to the small tusk of the holotype (USNM 488252) and has therefore been regarded as a female of O. peruvianus ( de Muizon et al., 1999) . In the following discussion, we do not exhaustively describe the new skull, but simply describe the new features O. peruvianus provided by the new specimen.
Holotype: USNM 488252, an incomplete skull lacking much of the left side and all of the auditory bones.
Referred specimens: SMNK PAL 2491, a relatively complete skull which lacks only the posterodorsal roof of the braincase, and which still retains the ear bones in situ; MNHN SAS 1613, a left periotic and partial tympanic; and MNHN SAS 1614, a left periotic.
Emended diagnosis: Odobenocetops peruvianus differs from O. leptodon in its snout, which is narrower at its apex and tapers dorsally (although the apex is still rounded); in the presence of large premaxillary foramina, anterior to the nares; in the absence of depressions for premaxillary sacs; in the shorter and shallower palate (especially in the anterior region); in the anterior border of the palate which is more Vshaped whereas it is more U-shaped in O. leptodon ; and in the deeply notched anterodorsal edge of the orbit, which is slightly concave in O. leptodon .
DESCRIPTION
TUSKS AND PREMAXILLAE
The major differences between the new specimen and the holotype are in the size of the tusks and morphology of the premaxillae. Both tusks are small and the left tusk is slightly smaller in diameter than the right. Neither of the tusks is complete. The left tusk is broken inside the sheath; however, the right tusk is only lacking its apex and, as preserved, is 23 cm long and is probably missing a few cm only. Therefore, the right tusk was very close to the length as well as the diameter of the small left tusk of O. leptodon . Although the tusk is not complete, given the length of the tusk (as preserved) and the sheath, it is certain that it was erupted. The premaxillary sheaths are small, and, in agreement with the sizes of the tusks, the left one is smaller than the right. Because of this slight asymmetry and although the apices of the left sheath and tusk are not preserved, it is likely that the left tusk was also erupted. Furthermore, the walls of the left sheath at its apex are very thin, which suggests that very little of the bone is missing. The apex of the right sheath is not completely preserved either, but given the thickness of the bone at the broken extremity, it is probable that no more than a few centimeters are lacking.
Basicondylar length: 412
Bizygomatic width: 158 ¥ 2 = 316e
Minimum width of the skull posterior to the postorbital processes: 116
The dorsal aspect of the premaxillae is very similar to that of the holotype although slightly more symmetrical. The posterior extension of the right prem axilla behind the nares is only slightly greater than that of the left, contrary to the condition of the holotype, in which the posterolateral process of the left premaxilla reaches the middle of the lateral edge of the left naris, whereas the right process almost reaches the posterior edge of the right naris. Since the premaxillae of SMNK PAL 2491 project posteriorly slightly behind the bony naris, this feature shows some variation within the species. The asymmetry of the tusks and alveolar processes may thus be reflected in the asymmetry of the posterior processes of the premaxillae. However, this inference must be made cautiously since most odontocetes (including monodontids) present an asymmetry of the dorsal bones of the skull, the right premaxilla and maxilla extending farther posteriorly than the left and slightly displacing the vertex on the left side of the skull. Nevertheless, the asymmetry of the male narwhal does not reflect the tusk asymmetry since the right maxilla is slightly more expanded posteriorly than the left, which carries the large tusk.
Minimum width between the dorsal edges of the orbits: 123
Maximum width between the postorbital processes: 226
Width of the snout between the antorbital notches: 87
Maximum width of both nares: 60
Maximum length of the right naris: 51.5
Width of the choanae: 96
Maximum width of the skull at the level of the exoccipitals: 250
Bicondylar width: 143
Width between the apices of the hamular processes: 128
Length of the lateral lobe of the left tympanic: 48.5
In SMNK PAL 2491 the snout is very complete and the structure of the bone is perfectly preserved. Therefore, muscle scars and foramina are clearly observable and provide important data. Anterior to the large premaxillary foramina is a rough, triangular surface, distinct on each bone, which reaches the apex of the snout. This area corresponds to the attachment scar for the nasal plug muscle in the other odontocetes. These muscle scars are so well marked that it is unlikely that there was no nasal plug muscle in O. peruvianus . It is therefore very probable that O. peruvianus had nasal plugs, which is to be expected in an odontocete, but was not clear on the holotype ( de Muizon et al., 2002). Posterior to the premaxillary foramina are two short saddle-shaped grooves, which form the anterior edges of the bony nares. The morphology of the new specimen confirms the conclusions of de Muizon et al. (2002) that, if premaxillary sacs (which are located between the premaxillary foramina and the anterior edge of the nares) were present in Odobenocetops peruvianus , they were very reduced, probably vestigial. Therefore, and considering the reduced space on the dorsal face of the rostrum, it is highly probable that O. peruvianus had no (or only a vestigial) melon. Although the nasal plugs, premaxillary sacs and melon are tightly related organs, the well-developed muscle scars for the insertion of the nasal plug muscle force us to conclude that O. peruvianus exhibited a strong reduction of the two latter organs but little reduction of the former.
On the anterior face of the snout the premaxillae bear several large foramina indicative of an extensive vascularization of the snout related to the inferred presence of a large and strong upper lip. The irregular surface of the bone in this region corresponds to the attachment of a very robust musculature of the lip. The morphology of the much better preserved female specimen of O. peruvianus (SMNK PAL 2491) confirms the hypotheses based on the holotype of the species.
MAXILLAE AND FRONTALS
Posterior to the nares, between the mesethmoid and the maxillae, are two small triangular medial portions of the frontals, which have the same condition as the maxillae in relation to the premaxillae at the anterior region of the nares. In other words, the maxillae, which lay on the frontals, have withdrawn posterior to the nares, uncovering the frontals. Posteriorly, the medial portions of the frontals become thin and progressively disappear after the maxillae become joined. Further posteriorly, the medial edges of the maxillae separate again and uncover a thin strip of the frontals which widens posteriorly. On the holotype it is probable that the lower portion of the mesethmoid posterior to (and overhung by) the Y-shaped portion which separates the nares (see de Muizon et al., 2002) is in fact the medial portion of the frontal observed on the female skull fused to the mesethmoid. A medial portion of the frontal is clearly present in O. leptodon (MNHN SAO 202). The extensions of the maxillae posterior to the nares narrow posteriorly but are, like the premaxillae and tusks, relatively symmetrical, whereas in the male the right maxilla is wider and extends further posteriorly than the left.
The preorbital process of the frontal of the female skull is anteriorly elongated and blade-like, as in the holotype. However, in this specimen it is clear that both the maxilla medially and the frontal laterally are components of the process. Therefore, although highly modified, the preorbital process of O. peruvianus resembles that of the other odontocetes in the combined participation of the maxilla and frontal. The postorbital process is similar in shape to that of the holotype but is larger and longer, indicating some individual variation of this character. A clear asymmetry of the frontal is present, as is observed in the other odontocetes, in which the right supraorbital process of the frontal is distinctly larger than the left.
PALATINE, PTERYGOID AND VOMER
The vomer is very wide, the maxillae do not participate in the construction of the palate, and the premaxillae and the palatines have a large V-shaped suture on both sides as is observed in the holotype. The posterior region of the palate is perfectly preserved and adds to the knowledge of the species. In the posterior region of the palate, the medial edges of the palatines are in contact, hiding the vomer for a few centimetres of its length. Further posteriorly the palatines separate again, uncovering the vomer ventrally. The median ridge of the vomer projects about 7 cm posterior to the posterior edges of the palatines. The sutures with the pterygoids are widely separated. The apices of the pterygoid hamuli are sharply pointed and extend toward the anterior angle of the basioccipital crests formed by the medial laminae of the pterygoids. They are almost in contact on the right side and the anterior end of the left medial lamina is apparently broken. Because of this projection and the long nonpalatal ridge of the vomer, the palate of O. peruvianus is clearly shorter than in O. leptodon and its posterior border is strongly concave. The pterygoid hamulus is transversely compressed and the lateral lamina of the pterygoid almost contacts the medial lamina. Therefore, the fossa for the pterygoid sinus is extremely reduced (as observed by de Muizon et al., 2002 on the holotype) and hardly present in O. peruvianus . It is clearly more reduced than in O. leptodon , in which the fossa is very small but conspicuous. As in O. leptodon , there is a short but continuous lateral lamina of the pterygoid, which contacts the alisphenoid, contrary to the condition in Recent delphinoids. However, in Recent monodontids, a continuous lateral lamina of the pterygoid, although almost absent from the pterygoid hamulus, is sometimes present and contacts the squamosal dorsally.
ALISPHENOID
The alisphenoid is similar to that of O. leptodon . It is very thick and bears an enormous foramen ovale not confluent with the cranial hiatus. de Muizon et al. (2002) stated that in the holotype of O. peruvianus the foramen ovale was confluent with the cranial hiatus. Because this is not the case in the three other skulls referred to Odobenocetops , it is possible that the condition observed in the holotype is due to a postmortem breakage of the alisphenoid. As in O. leptodon the external opening of the foramen ovale is formed anteriorly by the pterygoid and posteriorly by the alisphenoid. Internally, the foramen ovale is fully enclosed in the alisphenoid. It is noteworthy, however, that this condition is not easy to observe because of the partial fusion of the bones.
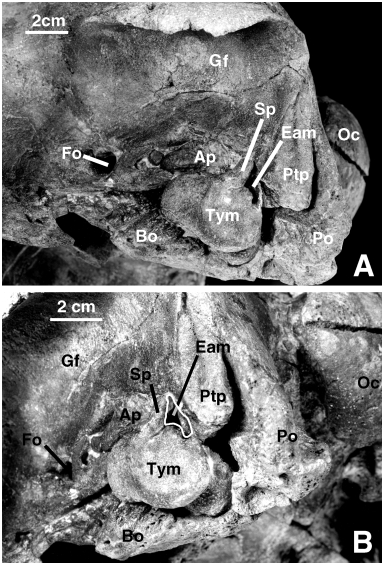
SQUAMOSAL AND EAR BONES
The squamosal of SMNK PAL 2491 is basically similar to that of the holotype. The tympanic and the periotic are preserved in situ on both sides of the specimen ( Fig. 15 View Figure 15 ). Since the bones could not be removed from the skull it is not possible to describe them thoroughly. However, it is clear that they are extremely similar to those referred to the species by de Muizon et al. (2002). The lateral face of the tympanic bears a large sigmoid process which is grossly perpendicular to the anteroposterior axis of the bone and not shifted posteriorly as in the classical (i.e. non-odobenocetopsid) delphinoids. The apex of the process is straight, rounded, and not strongly recurved anteriorly as is observed in the classical delphinoids. Posterior to it is a low conical process and a relatively large anterior border of the posterior process. These three elements surround the external auditory meatus and, as a consequence of their morphology, the meatus is much larger than in the classical delphinoids. It is slightly smaller than in O. leptodon (SMNK PAL 2492) . This condition would indicate a more efficient (effective) tympanic membrane in the Odobenocetopsidae and therefore possibly better sound perception via the middle ear than in the other odontocetes.
The lateral wall of the tympanic is high, as is observed in O. leptodon . This condition indicates that the middle ear cavity was larger than in the classical delphinoids. It resembles the condition of most early diverging odontocetes ( Squalodontidae , Squalodelphinidae , Platanistidae , Eurhinodelphinidae ), and could represent a plesiomorphic condition or, more likely, a reversal corresponding to a need for enlargement of the middle ear cavity. The anterior extremity of the tympanic is blunt and rounded, as is observed in the other delphinoids.
The ventral lobes are well separated by a deep median groove which is well-marked on the posterior two-thirds of the bone. Their posterior extremities are rounded and wide and the medial lobe is slightly posterior to the lateral. This condition is similar to that of O. leptodon but clearly differs from that of the Recent monodontids, in which the posterior extremity of the medial lobe is narrower and more anterior than that of the lateral lobe. This feature is especially obvious in Monodon . It is also present to a lesser extent in the delphinids and phocoenids. Again, the odobenocetopsid condition is found in the early diverging odontocetes mentioned above, but is regarded here as a reversal (rather than a plesiomorphy).
Not much can be said about the periotic, which is preserved in situ, except that it has the massive morphology observed on the referred specimens described by de Muizon et al. (2002). Contrary to what is observed in the classical delphinoids, the lateral portion of the bone is closely appressed against the squamosal, in this respect resembling the condition observed in the platanistoids (sensu de Muizon, 1987; 1991; 1994; Fordyce & Barnes, 1994; Fordyce et al., 1994; Fordyce, 1994; not sensu McKenna & Bell, 1997). Furthermore, the posterior process of the periotic is deeply wedged between the squamosal and the exoccipital, which is in agreement with the long posterior process observed on MNHN SAS 1614 (an isolated periotic from the same horizon as the holotype of O. peruvianus and referred to this species). A similar condition is present in Delphinapterus ( Kasuya, 1973) and Monodon . The delphinids and phocoenids are characterized by a petrotympanic completely isolated from the skull and maintained in situ by ligaments only ( Fraser & Purves, 1960). Early derived odontocetes such as platanistoids have a tympanic and a periotic still articulated with the squamosal, which is regarded as a plesiomophic character state ( de Muizon, 1987; 1991; 1994). Therefore, the condition of Odobenocetops and the Monodontidae could be regarded as plesiomorphic. However, because they have synapomorphies of the Delphinoidea absent in the Platanistoidea, we rather consider the condition of Odobenocetops as a reversal due to its hyperspecialization. The fact that a condition approaching this one is also found in monodontids could represent a synapomorphy of the two families, possibly related to sound transmission.
In general, the periotic and tympanic of Odobenocetops have relatively tight and extensive contacts with the surrounding bones: basioccipital and squamosal for the tympanic, and squamosal and exoccipital for the periotic. The lateral tuberosity and rim of the anterior process of the periotic are closely appressed against the squamosal in a similar pattern to that observed in the platanistoids. The squamosal in this region is greatly thickened and forms the medial border of the trough-like glenoid cavity.
On the anterior region of the anterior process of the periotic are several independent bony nodules which are appressed against the anterior process and apparently part of it. Such a condition has been observed in O. leptodon and is also present sometimes in Platanista .
BASIOCCIPITAL
The basioccipital is extremely wide and basically similar to that of O. leptodon although slightly wider. The basioccipital crests are large and undergird the tympanics, which are partially imbedded in their lateral face. The posteromedial faces of the basioccipital crests bear very large and deep muscular scars for insertion of the scalenus, at least as strong as in O. leptodon .
EXOCCIPITAL
The paroccipital process is very large and stout. It differs from that of the holotype in being orientated ventrally and not ventrolaterally. Its ventromedial angle contacts the posteroventral angle of the basioccipital crest and encloses the jugular notch to form a relatively large window, as is observed in O. leptodon . The exoccipital border of the window is very thick and forms an elevated crest. This indicates a strong scalenus dorsalis, which is inserted on the posteroventral face of the paroccipital process. This part of the occipital indicates an exceptionally strong musculature of the neck as was noted in regard to the holotype of Odobenocetops peruvianus ( de Muizon et al., 2002) . The anterior face of the paroccipital process does not present any fossa for the posterior sinus such as is observed in many other delphinoids. Fraser & Purves (1960) have stated that the absence of a fossa is not equivalent to absence of a posterior sinus. As a matter of fact, in the Recent monodontids the anterior face of the paroccipital process is not excavated by a fossa and a small posterior sinus is present. However, the presence of a large sinus is related to a deep fossa. In SMNK PAL 2491 the anterior face of the paroccipital process has a reduced area and is not indicative of a large posterior sinus. This statement contrasts with the observations made on the holotype of O. peruvianus ( de Muizon et al., 2002) .
The occipital condyles are more convex and salient than in any recent delphinoid and confirm the great mobility of the neck which was noted with regard to the holotype. In fact, the occipital condyles form a quarter of a sphere ( Figs 13B,D View Figure 13 and 14 View Figure 14 ) and are proportionally larger and more convex than in any other cetacean (Recent or fossil) or in the walrus. The morphology of the condyles of SMNK PAL 2491 therefore indicates an exceptional mobility of the neck in Odobenocetops ; certainly much greater than that of any other odontocete.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.